Class Aves
Order Galliformes
Family Phasianidae
Subfamily Tetraoninae
Grouse—Grouse // Centrocercus urophasianus/minimus—Greater/Gunnison Sage-Grouse // Dendragapus obscurus—Dusky Grouse // Lagopus—Ptarmigan // Tympanuchus sp.—Prairie-chickens // Tympanuchus pallidicinctus—Lesser Prairie-chickens
Members of the subfamily are largely ground dwellers whose flight usually is limited to relatively short distances. They mostly are equipped with short, stout claws for scratching in the soil for food, although the Greater Sage-Grouse feeds largely on herbage (Ligon 1961).
Literature. Ligon 1961.
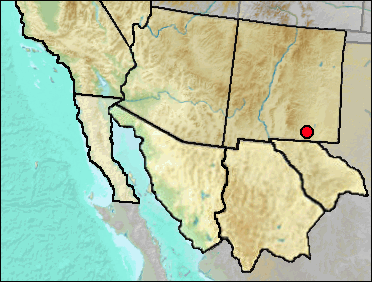
Howard (1971:238) listed without comment "Grouse, 2 spp.?" for Dark Canyon Cave. Lacking further information, these are merely listed by this common name.
Sites.
Mid/Late Wisconsin: Dark Canyon Cave (Howard 1971).
Literature. Howard 1971.
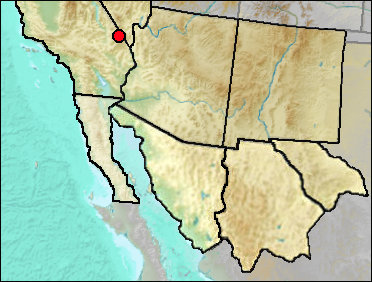
Centrocercus records are as given in the published records. However, since C. minimus has only recently been described, the possibility is that some earlier records actually may pertain to that species.
Sites.
Late Wisconsin/Holocene: Kokoweef Cave (Reynolds, Reynolds, et al. 1991: ? gen.).
Literature. Reynolds, Reynolds, et al. 1991.
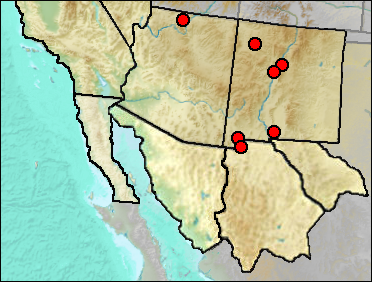
 Fig. 1. Centrocercus urophasianus. Photograph by Dave Menke,
courtesy of the US Fish & Wildlife Service.
Fig. 1. Centrocercus urophasianus. Photograph by Dave Menke,
courtesy of the US Fish & Wildlife Service.
Until 2000, only one species of sage-grouse was recognized. In that year, however, Young et al. described a second species, Centrocercus minimus, inhabiting southwestern Colorado and adjacent southeastern Utah. They also speculated that the now extirpated sage-grouse of New Mexico was this species. Centrocercus minimus was described as having a body mass about 27-33% less than that of the Greater Sage-Grouse, with males in the upper portion of the range and females in the lower.
At present, the only skeletal comparative material in the UTEP bird collection is a female from Wyoming. Without data on skeletal size differences between the sexes, it is not clear which species is represented by the UTEP fossils. The Isleta Cave material appears to be very slightly larger than the modern female; the U-bar material may represent male and female remains. To summarize, it is unclear which species is represented, but the limited data available suggest the Greater Sage-Grouse is represented in these two sites. In the absence of size information on the other occurrences of sage-grouse in Arizona and New Mexico, they are identified merely as one of these two species..
The following information on modern populations of the Greater Sage-Grouse is primarily from Johnsgard (1973). Various species of sagebrush (Artemisia) play a major role in modern Sage Grouse ecology, with the big sagebrushes (A. tridentata and similar species) being vital. The majority of nest sites occur in sagebrush stands, and sagebrush is the major food item. Sagebrush in wintering grounds (which may be 30 or more miles distant from summer habitat) provides nearly 100% of winter food and significant amounts at other times of year. Forbs and grasses provide most other food items in the adult, resulting in the preferred habitat being a sagebrush-grassland.
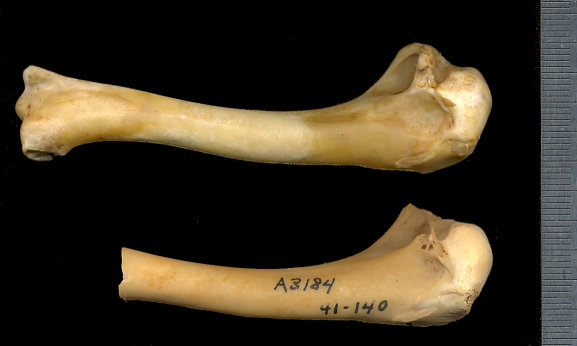
Fig. 2. Centrocercus urophasianus right humerus. Modern specimen (UTEP 2233) at top; specimen of Centrocercus cf. urophasianus from Isleta Cave No. 1 at bottom. UTEP 41-140. Metric scale.
"At one time this species was found virtually wherever sagebrush (Artemisia, especially A. tridentata) occurred, throughout many of the western and intermontane states" (Johnsgard 1973:157). Northern New Mexican sage-grouse were exterminated historically. The New Mexico Ornithological Society checklist of New Mexican birds lists the former inhabitants of the state as C. minimus.
The life history data strongly uphold the presence of significant stands of A. tridentata-type sagebrush in the western half of New Mexico during the late Wisconsin. Although other data suggest sagebrush also in the southeastern portions of the state, presence of the grouse has not been recognized. Brodkorb (1964) reported it from Rocky Arroyo (= Burnet Cave), but confused a statement by Howard and Miller (1933) that contrasted its presence at Conkling and Shelter caves with its absence at Rocky Arroyo.
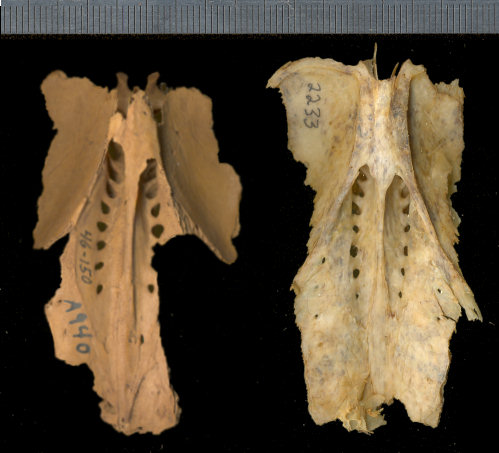 Fig.
3. Centrocercus urophasianus synsacrum and partial ilia. Fossil specimen from
Isleta Cave No. 2 at left (UTEP 46-150); modern specimen at right. Metric
scale.
Fig.
3. Centrocercus urophasianus synsacrum and partial ilia. Fossil specimen from
Isleta Cave No. 2 at left (UTEP 46-150); modern specimen at right. Metric
scale.
Sites.
Mid/Late Wisconsin: Shelter Cave (Howard and Miller 1933).
Mid Wisconsin: U-Bar Cave (UTEP).
Late Wisconsin: Sandia Cave (Brasso and Emslie 2006); Sheep Camp Shelter (Gillespie 1985); U-Bar Cave 14-15 ka (Harris 1989); U-Bar Cave 15-18 ka (Harris 1989).
Late Wisconsin/Holocene: Conkling Cavern (Howard and Miller 1933); Isleta Cave No. 1 (Harris 1993c); Isleta Cave No. 2 (Harris 1993c); Howell's Ridge Cave (Howard 1962); Stanton's Cave (Rea and Hargrave 1984).
Literature. Brasso and Emslie 2006; Brodkorb 1964; Gillespie 1985; Harris 1989, Harris 1993c; Howard 1962, Howard and Miller 1933; Johnsgard 1973; Rea and Hargrave 1984; Young et al. 2000.
The Dusky Grouse was the most common bird recovered from Sandia Cave, with 174 identified bones represented a minimum of 24 individuals (Brasso and Emslie 2006). It was represented by a single element in the much smaller avifauna from Marmot Cave.
Johnsgard (1973) reports that the geographic range of the Dusky Grouse closely conforms to the distribution of true firs (Abies) and Douglas Fir (Pseudotsuga). Seasonal changes in elevation are common, with reproduction tending to occur at lower elevations and wintering near or above timberline. Modern occurrences in our region are in the higher mountains of northern and western New Mexico; the total range extends north into the Yukon.
Sites.
Late Wisconsin: Marmot Cave (Brasso and Emslie 2006); Sandia Cave (Brasso and Emslie 2006).
Literature. Brasso and Emslie 2006; Johnsgard 1973.
 Fig. 1. Willow Ptarmigan (Lagopus lagopus). Photograph by Tim
Bowman, courtesy of the US Fish & Wildlife Service.
Fig. 1. Willow Ptarmigan (Lagopus lagopus). Photograph by Tim
Bowman, courtesy of the US Fish & Wildlife Service.
Ptarmigans are birds of arctic tundra or, in the western U.S., above-timberline habitats (Johnsgard 1973). Today, the North American species are limited to north of the U.S./Canada border except for the White-tailed Ptarmigan (Lagopus leucura). This taxon approaches our area in those high mountains of northern New Mexico that extend above timberline; presumably it is this species represented in Sandia Cave. However, although Brasso and Emslie (2006) state that they cannot rule out a large, temporal form of this species, they note that in size and characters the fossil is similar to those of the other two North American representative of the genus, the Willow Ptarmigan and the Rock Ptarmigan.
Sites.
Late Wisconsin: Sandia Cave (Brasso and Emslie 2006).
Literature. Brasso and Emslie 2006; Johnsgard 1973.
As the name implies, prairie chickens are predominantly grassland animals. The historic species of the eastern portions of our region was T. pallidicinctus, which still holds on with a reduced geographic range in parts of eastern New Mexico and adjacent Texas. The original range extended through western Oklahoma and western Kansas; the Greater Prairie-Chicken (T. cupido) replaces T. pallidicinctus to the east and north. The geographic range of the Sharp-Tailed Grouse (T. phasianellus) reaches south to extreme northeastern New Mexico (Johnsgard 1973).
Harris (1993c) reported cf. Tympanuchus from Harris' Pocket. The specimen, the distal 4/5 of an ulna, is stockier than the UTEP limited comparative material of T. pallidicinctus and T. cupido, but appears less so than that of the one modern female specimen of Centrocercus urophasianus available. Our comparative material is insufficient to judge the degree of sexual size dimorphism in Tympanuchus. The status of this specimen is changed to "? Tympanuchus."
Sites.
Late Wisconsin: Harris' Pocket (UTEP: ?).
Literature. Harris 1993c; Johnsgard 1973.
 Fig. 1. Lesser Prairie-chicken. U.S. Fish & Wildlife Service
photograph.
Fig. 1. Lesser Prairie-chicken. U.S. Fish & Wildlife Service
photograph.
The preferred habitat of the Lesser Prairie-Chicken is sand-sage grasslands (Artemisia filifolia-Andropogon) and shin-oak grasslands (Quercus havardii-Andropogon), both of which are associated with notably sandy substrates (Johnsgard 1973). In New Mexico, this means the extreme eastern portions of the state, east of the Pecos.
In view of the unlikelihood of extensive patches of sandy substratum in the immediate vicinity of the southeastern New Mexican sites, one possibility is that a similar plant-structured, but non-sandy, habitat was utilized during the Pleistocene; conversely, the remains may have been brought to the sites by predators.
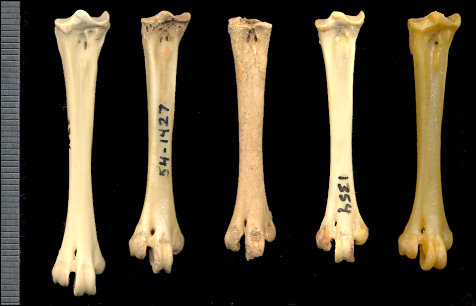 Fig. 2. Comparison
of the tarsometarsi of T. cupido (left), fossils from TT II (54-1427) and NW
Talus Slope (22-358), and two modern specimens of T. pallidicinctus
(right).
Fig. 2. Comparison
of the tarsometarsi of T. cupido (left), fossils from TT II (54-1427) and NW
Talus Slope (22-358), and two modern specimens of T. pallidicinctus
(right).
Sites.
Mid Wisconsin: Pendejo Cave (Harris 1993c).
Mid/Late Wisconsin: Dark Canyon Cave (Howard 1971: ?; UTEP); Hampton Court (Harris 1993c); NW Talus Slope (UTEP).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Animal Fair 18-20 ka (Harris 1989: cf.); Big Manhole Cave (Harris 1993c); Charlies Parlor (Harris 1989: cf.); Stalag 17 (Harris 1993c); TT II (Harris 1993c).
Late Wisconsin/Holocene: Burnet Cave (Wetmore 1932).
Literature. Harris 1989, 1993c; Howard 1971; Johnsgard 1973; Wetmore 1932.
Last Update: 11 Jun 2014