Class Mammalia
Order Artiodactyla
Suborder Ruminantia
Family Antilocapridae
Synonyms. Breameryx arizonensis.
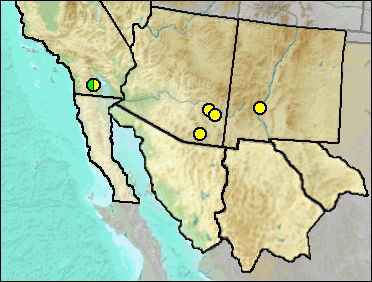
Capromeryx is recognized in the Pleistocene Blancan of New Mexico by a partial calcaneum (Morgan et al. 2011). The larger size separates it from the late Pleistocene C. furcifer, and the only small antilocaprid known from the Southwestern late Blancan is this species (Morgan et al. 2011).
Sites.
Late Blancan: 111 Ranch (Morgan and White 2005); Caballo (Morgan et al. 2011: ?); Curtis Ranch (Morgan and White); San Simon Fauna (Morgan and White 2005).
Irvingtonian: Anza-Borrego (Murray 2006).
Literature. Morgan and White 2005; Morgan et al. 2011; Murray2006.
Synonyms. Capromeryx minor, Capromeryx mexicana, Capromeryx minimus, Breameryx minor.
Several species of Capromeryx have been described, but the two-pronged horn cores generally have been considered necessary for sure identification to species. Despite a number of recorded specimens, only three records from our region appear to be based on horn core material (Pendejo Cave, Shelter Cave, and Tramperos Creek, though Doolittle Cave has the base of a horn core). Lacking horn cores, most fossil elements have been assigned to the genus on the basis of size (see, for example, Fig. 2 of the Stockoceros account), but in the absence of horn cores, generally no determination of species has been made in the literature. However, White and Morgan (2011) considered, pending formal taxonomic revision, all Capromeryx with the anterior prong of the horn core less than 50% of the length of the posterior to be assignable to C. furcifer; this would include all Rancholabrean and probably all Irvingtonian specimens for which horn cores are available, indicating that likely all Pleistocene, post-Blancan Capromeryx are assignable to C. furcifer. Following this reasoning, all Capromeryx considered here are assigned to that species.

Fig. 1. Skeleton of Capromeryx furcifer, Rancho La Brea. Photograph by David Monniaux, licensed under Creative Commons Attribution-Share Alike 3.0 Unported

Fig. 2. Three views of the horn cores of Capromeryx furcifer. The small structure is the anterior prong; the dorsal end is missing. A straight groove or furrow runs up the posterior prong directly behind the anterior prong and another on the anteromedial side. Pendejo Cave specimen (9366.G.989.337.1). Scale in mm.
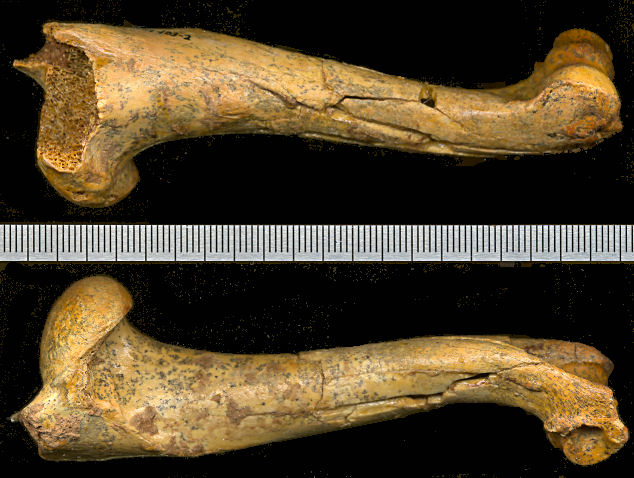
Fig. 3. Medial and lateral views of the left humerus of Capromeryx (5689-103-2) from U-Bar Cave. Scale in mm.
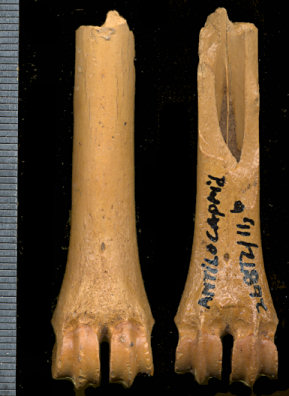
Fig. 4. Anterior and posterior views of the distal metacarpal of Capromeryx from U-Bar Cave (USNM 26872/11, b). Scale in mm.
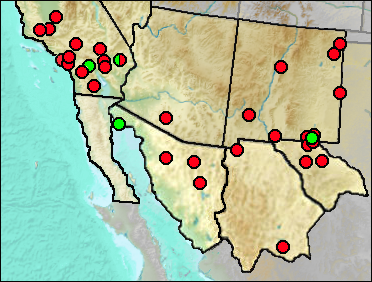
Sites.
Pleistocene: Perico Creek (Morgan and Lucas 2005).
Irvingtonian: Bautista Badlands (Frick 1921); El Golfo (Croxen et al. 2007).
Medial Irvingtonian: Slaughter Canyon Cave (Morgan and Harris 2015).
Irvingtonian/Rancholabrean: Cadiz (Jefferson 2014: ?).
Rancholabrean: Bow Willow (Jefferson 2014); Campbell Hill (Jefferson 1991b); Centinella Park (Jefferson 1991b); Chino Hills (Jefferson 2014); Eagle Crest (Jefferson 2014); Harbor Freeway, Athens (Jefferson 1991b); La Playa (White et al. 2010); Los Angeles Police Station (Jefferson 1991b: cf. gen.); Ludlow Cave (Jefferson 1991b); Pinto Basin (Jefferson 2014: Cf. gen.); Tramperos Creek (White and Morgan 2011); Térapa (White et al. 2010).
Sangamon: La Brisca (Van Devender et al. 1985: cf.); San Pedro Lumber Co. (Jefferson 1991b: cf.).
Wisconsin: Costeau Pit (Jefferson 1991b); Doolittle Cave (Cosgrove 1947).
Early/Early-Mid Wisconsin: Lost Valley (Harris 1993c); Rm Vanishing Floor (Harris 1993c); Sabertooth Camel Maze (Harris 1993c).
Mid Wisconsin: McKittrick (Schultz 1937); Pendejo Cave (Harris 2003); U-Bar Cave (Harris 1987).
Mid/Late Wisconsin: Animal Fair (Harris 1993c); Big Manhole Cave (this work); Diamond Valley (Springer et al. 2009); Hampton Court (Harris 1993c); Pit N&W Animal Fair (Harris 1993c); Rancho La Brea (Stock and Harris 1992).
Mid/Late Wisconsin/Holocene: Jimenez Cave (Messing 1986); Sierra Diablo Cave (UTEP).
Mid Wisconsin/Holocene: Shelter Cave (Stock 1932).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Big Manhole Cave (UTEP); Blackwater Draw Fauna (Slaughter 1975); Conkling Cavern (Conkling 1932); Dust Cave (Harris and Hearst 2012: cf.); Fowlkes Cave (Dalquest and Stangl 1984b); Human Corridor (Harris 1993c); Maricopa (Jefferson 1991b); Muskox Cave (White and Morgan 2011); Pendejo Cave (Harris 2003); Sandia Cave (Thompson and Morgan 2001); Schuiling Cave (Jefferson 1991b); Stalag 17 (Harris 1993c: cf.); TT II (Harris 1993c: cf.); U-Bar Cave 14-15 ka (Harris 1989); U-Bar Cave 15-18 (Harris 1989: ?); U-Bar Cave 18-20 ka (Harris 1989); Ventana Cave (Mead et al. 2005).
Changes.
Mid/Late Wisconsin: Dark Canyon Cave (Harris 1993c) re-identified as Stockoceros conklingi.
Late Wisconsin: Balcony Room (Harris 1993c: cf.) re-identified as Stockoceros conklingi.
Literature.
Conkling 1932; Croxen et al. 2007; Cosgrove 1947; Dalquest and Stangl 1984b; Frick 1921; Harris 1987, 1989, 1993c, 2003; Harris and Hearst 2012; Jefferson 1991b, 2014; Mead et al. 2005; Morgan 2003; Morgan and Harris 2015; Morgan and Lucas 2005; Schultz 1937; Slaughter 1975; Springer et al. 2009); Stock 1932; Stock and Harris 1992); Thompson and Morgan 2001; Van Devender et al. 1985; White and Morgan 2011.
Last Update: 4 Feb 2016