Class Mammalia
Order Rodentia
Family Heteromyidae
Dipodomys sp.—Kangaroo Rats // Dipodomys small—Small Kangaroo Rat // Dipodomys agilis—Agile Kangaroo Rat // Dipodomys compactus—Gulf Coast Kangaroo Rat // Dipodomys deserti—Desert Kangaroo Rat // Dipodomys gidleyi—Gidley's Kangaroo Rat // Dipodomys hibbardi—Hibbard's Kangaroo Rat // Dipodomys merriami—Merriam's Kangaroo Rat // Dipodomys merriami/ordii—Merriam's or Ord's Kangaroo Rat // Dipodomys microps—Chisel-toothed Kangaroo Rat // Dipodomys ordii—Ord's Kangaroo Rat // Dipodomys nelsoni/spectabilis—Nelson's or Banner-tailed Kangaroo Rat // Dipodomys panamintinus—Panamint Kangaroo Rat // Dipodomys spectabilis—Banner-tailed Kangaroo Rat
Kangaroo rats received their name because of similarities of locomotion and the morphology that goes with it between these rodents and the marsupials. The hind limbs are massively developed for the size of the animal and the forelimbs diminutive. Other characteristics include huge feet, a very long tail for balance, and an oversized head, thanks to the greatly enlarged bullae.
Kangaroo rats tend to forage in open spaces, using their paws to stuff seeds into the cheek pouches and, with the food safely stored, then retreating to cover. While in the open, they are subject to predation, especially from owls. Owls have feathers adapted for silent flight, but checking their dive upon potential food produces a low frequency sound. This sound apparently is not detected by most rodents. However, laboratory studies indicate that the large bullae of kangaroo rats are specifically adapted to pick up the slight sound made a split second before the rat is in the grip of the owl's talons. In that brief window of time, the rat undertakes a panic jump that may (or may not) save it. Since kangaroo remains tend to be plentiful in owl pellets, it can be assumed that selective pressure for this hearing is strong.
Presence of kangaroo rats in our region is taken to indicate presence of open areas basically free of vegetation (though with bushes or other vegetation nearby for cover).
D. merriami and D. ordii are similar in size (though D. ordii is a bit larger) and may end up identified merely as "Dipodomys" or as "Dipodomys small" (to separate them from D. spectabilis, which is large enough as an adult to be identified by size alone in the eastern portion of the region). Identifications to modern species may break down in early sites, since extinct or extralimital species may be present.
Kangaroo rats construct burrows for protection, usually with several openings to the burrow system. As with their relatives, the pocket gophers, they tend to be antisocial except during the breeding season, but two or three species may occur sympatrically.
Dalquest and Stangl (1984b) found that the lower incisor breadth is greater than 1.0 mm in D. spectabilis, 0.8 to 0.95 mm in D. ordii, and less than 0.8 mm in D. merriami. They also found that the alveolar tooth row length would separate the three species, with tooth rows of D. spectabilis greater than 5.6 mm, those of D. ordii 4.8-5.5 mm, and those of D. merriami less than 4.75 mm. The incisor breadth is most useful, since the posterior rim of bone adjacent to the alveolus of m3 often is broken away in fossils.
Literature. Dalquest and Stangl 1984b.
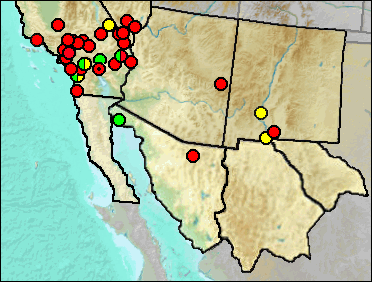
Sites.
Identification only to the generic level likely means that the record is based on material that cannot be identified to a lower taxonomic level or that the species has not been recorded in the literature. The La Brisca specimens are within the size range of D. ordii and D. merriami (Van Devender et al. 1985).
Sites.
Late Blancan: Caballo (Morgan et al. 2011); La Union (Morgan and Lucas 2003); Jack Rabbit Trail, San Timoteo Badlands (Albright 2000); Tecopa Lake Beds (Woodburne and Whistler 1991: two new spp.).
Late Blancan/Early Irvingtonian: Elsinore: Mimomys (Pajak et al. 1996).
Irvingtonian: El Casco, San Timoteo Badlands (Albright 2000); El Golfo (Croxen et al. 2007); Elsinore: Microtus/Mammuthus (Pajak et al. 1996).
Early Irvingtonian: Gypsum Ridge (Wagner and Prothero 2001: cf.).
Late Irvingtonian: Elsinore: Pauba Formation [large and small species] (Pajak et al. 1996).
Late Irvingtonian/Early Rancholabrean: Cadiz (Jefferson 2014); Thorn, Victorville (Jefferson 1991b).
Rancholabrean: Bedford Properties (Jefferson 1991b); Centennial Parkway, Las Vegas Valley (Jefferson et al. 2015: two species); Chino Hills (Jefferson 2014); Clay Mine Road (Jefferson 2014); Cool Water Coal Gasification Solid Waste Site (Jefferson 1991b); Detention Basin, Upper Las Vegas Wash (Jefferson et al. 2015: large and small species); Eagle Crest (Jefferson 2014: large and small species); Edwards Air Force Base (Jefferson 2014); Glen Abbey, Bonita (Jefferson 1991b); Hawes (Jefferson 2014); Helendale (Jefferson 1991a); Hoffman Road (Jefferson 2014); Lake View Hot Springs (Jefferson 2014); Park La Brea A (Jefferson 2014); Piute Ponds (Jefferson 2014); Piute Valley (Jefferson 1991b); Silver Lake (Jefferson 1991b); Wilshire Blvd., 10580 The Wilshire, Los Angeles (Jefferson 1991b).
Rancholabrean/Early Holocene: Metro Rail Universal City Station (Jefferson 2014).
Sangamon: Kelly Pits (Jefferson 1991b); La Brisca (Van Devender et al. 1985); Naval Fuel Reserve Quarry (Jefferson 1991b).
Wisconsin: Carpinteria (Wilson 1933); Costeau Pit (Jefferson 1991b); Danby Lake (Jefferson 1991c).
Mid Wisconsin: Devil Peak (Reynolds, Reynolds, and Bell 1991); Screaming Neotoma Cave (Glennon 1994).
Mid/Late Wisconsin: Diamond Valley (Springer et al. 2009); Rampart Cave (Lindsay and Tessman 1974).
Late Wisconsin: Antelope Cave (Reynolds, Reynolds, Bell, and Pitzer 1991); Falling Arches (Jefferson 1991b); Glendale (Jefferson et al. 2015); New Water Mountains (Mead et al. 2005); Red Tail Peak Midden (Jefferson 1991b); Tsuma Properties, San Clemente (Jefferson 2014); Tunnel Ridge Midden (Jefferson 1991b); Valley Wells (Springer et al. 2010).
Rancholabrean/Holocene: Arvin Landfill (Jefferson 2014).
Late Wisconsin/Holocene: Calico Lakes (Jefferson 1991b); Conkling Cavern (Harris 1993c); Kokoweef Cave (Reynolds, Reynolds, et al. 1991); Luz Foundation (Jefferson 1991b); Luz Solar Trough (Jefferson 1991b); Newberry Cave (Jefferson 1991b); Schuiling Cave (Jefferson 1991b); Solar One (Jefferson 1991b); Tule Springs (Mawby 1967).
Literature. Albright 2000; Croxen et al. 2007; Glennon 1994; Harris 1993c; Jefferson 1991b, 1991c, 2014; Jefferson et al. 2015; Lindsay and Tessman 1974; Mawby 1967; Mead et al. 2005; Morgan and Lucas 2003; Morgan et al. 2011; Pajak et al. 1996; Reynolds, Reynolds, and Bell 1991; Reynolds, Reynolds, et al. 1991; Reynolds, Reynolds, Bell, and Pitzer 1991; Springer et al. 2009, 2010; Van Devender et al. 1985; Wagner and Prothero 2001; Wilson 1933; Woodburne and Whistler 1991.
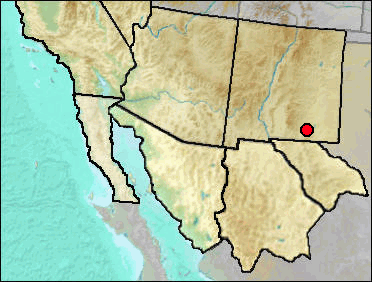
As noted above, presumably either D. merriami or D. ordii is represented.
Sites.
Early/Early-Mid Wisconsin: Rm Vanishing Floor; Sabertooth Camel Maze (Harris 1993c).
Literature. Harris 1993c.
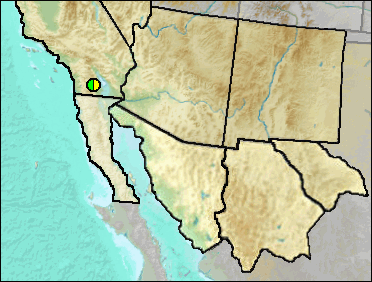
According to Murray (2008), a diagnosis was included in a M.S. thesis by Cunningham (1984), but has not formally named nor published.
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Cunningham 1984; Murray 2008.
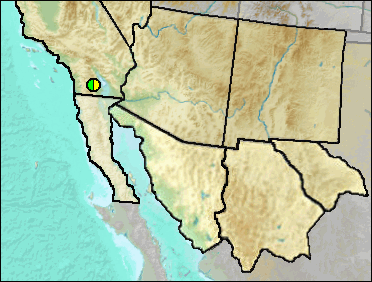
According to Murray (2008), a diagnosis was included in a M.S. thesis by Cunningham (1984), but has not been formally named nor published.
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Cunningham 1984; Murray 2008.
Sites.
Sangamon: Naval Housing Unit (Jefferson 1991b: cf.); Newport Bay Mesa (Jefferson 1991b: cf.).
Mid/Late Wisconsin: Rancho La Brea (Stock and Harris.
Mid Wisconsin: Pacific City (Wake and Roeder 2009: cf.).
Late Wisconsin: Maricopa (Jefferson 1991b).
Literature. Jefferson 1991b; Stock and Harris 1992; Wake and Roeder 2009.
One is strongly inclined to assume the identification is incorrect—the current range is limited to the Gulf of Mexico coastal areas of Texas and south along the Mexican coast. The assignment of fossils to this taxon appears limited to California (fossilworks, accessed 4 July 2014). Zakrzewski (1981) noted that D. compactus may be considered as the most primitive of extant species. This being the case, it seems probable that the name is being applied to a morphological grade rather than a biological species.
Sites.
Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Murray 2008; Zakrzewski 1981.
The Desert Kangaroo Rat is one of the larger of the genus. Distribution today is the western Great Basin and south into the Sonoran Desert of southwestern Arizona and northern Mexico.
Sites.
Rancholabrean: Saltmarsh (Jefferson 1991b); Siphon 11 (Jefferson 1991b).
Late Wisconsin: Mountain View Country Club (Jefferson 2014: cf.); Pintwater Cave (Hockett 2000).
Literature. Hockett 2000; Jefferson 1991b, 2014.
Sites.
Late Blancan: 111 Ranch (Morgan and White 2005); Curtis Ranch (Morgan and White 2005).
Literature. Morgan and White 2005.
Dipodomys hibbardi was described by Zakrzewski (1981) as approximately evolutionarily intermediate between Prodipodomys and the living Dipodomys compactus, with shorter dentine tracts than the living form, but higher than in Prodipodomys. He also pointed out that root development was intermediate.
Sites.
Late Blancan: 111 Ranch (Morgan and White 2005); Anza-Borrego (Murray 2008); San Simon Fauna (Morgan and White 2005).
Literature. Morgan and White 2005; Murray 2008; Zakrzewski 1981.
This large kangaroo rat currently has a very limited range in central California.
Sites.
Mid Wisconsin: McKittrick (Schultz 1937: cf.).
Literature. Schultz 1937.
Merriam's Kangaroo Rat is an arid-land species essentially limited to Lower Sonoran habitats. It tends to prefer firmer soils than Ord's Kangaroo Rat. However, there may be overlap in habitat usage, and at times both of these smaller species may be taken in the same trap line.
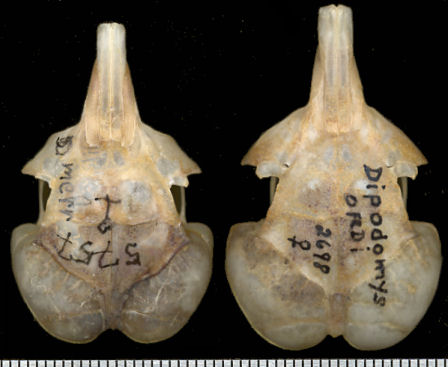 Aside from the discriminatory measurements
recorded in the generic account, the auditory bullae tend to be somewhat differently developed than
in D. ordii. This and other subtle differences can be seen in Fig. 1.
Aside from the discriminatory measurements
recorded in the generic account, the auditory bullae tend to be somewhat differently developed than
in D. ordii. This and other subtle differences can be seen in Fig. 1.
Fig. 1. Comparison of the skulls of D. merriami (left) and D. ordii. Scale is in mm.
Although reported from the Isleta Caves (1993c), it does not seem to be vouchered and is rejected.
Sites.
Rancholabrean: Bitter Springs Playa (Jefferson 2014: cf.);Saltmarsh (Jefferson 1991b).
Rancholabrean/?Early Holocene: Mitchell Caverns (Jefferson 1991b: aff.).
Mid Wisconsin: Castle Mountains (Mead et al. 2005).
Late Wisconsin: Mountain View Country Club (Jefferson 2014: cf.); Murray Springs (Mead et al. 2005); Wellton Hills (Mead et al. 2005: cf.); Wolcott Peak (Mead et al. 2005).
Mid/Late Wisconsin/Holocene: Pintwater Cave (Jefferson et al. 2015).
Late Wisconsin/Holocene: Fowlkes Cave (Dalquest and Stangl 1984b); Howell's Ridge Cave (Harris 1993c); Pendejo Cave (Harris 2003)
Late Wisconsin/Holocene: Isleta Caves (Harris 1993c), rejected.
Literature. Dalquest and
Stangl 1984b; Harris 1993c, 2003; Jefferson 1991b, 2014; Jefferson et al. 2015; Mead et al. 2005.
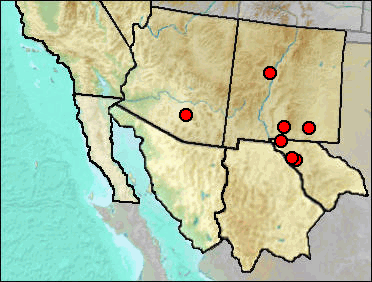
The material on which the following records are based are of a nature not allowing discrimination between the two species.
Sites.
Mid Wisconsin: Tank Trap Wash (Van Devender et al. 1987).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Balcony Room (UTEP); Dry Cave <13 ka (Harris 1993c); Navar Ranch (Van Devender et al. 1987); Pendejo Cave (Harris 2003); Picacho Peak (Van Devender et al. 1991); Steeruwitz Hills #1(3) (Van Devender and Bradley 1990).
Wisconsin/Holocene: Isleta Cave No. 2 (UTEP).
Literature. Harris 1993c, 2003; Van Devender and Bradley 1990; Van Devender et al. 1987, 1991.
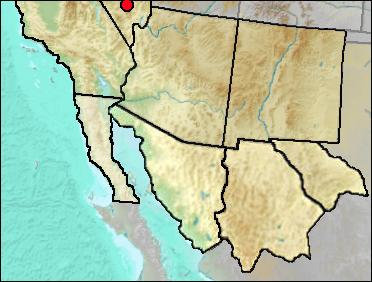
This is a Great Basin form today.
Sites.
Late Wisconsin: Pintwater Cave (Hockett 2000).
Literature. Hockett 2000.
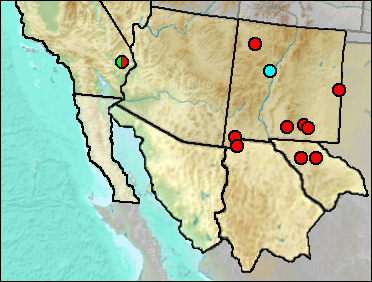
Unlike D. merriami, which has a southern distribution, D. ordii ranges in the West from southern Canada deep into Mexico. It prefers sandy soils, but may occur in various aspects of grassland and shrubby habitats. It only marginally ranges as high as woodland.
Synonyms. Perodipus montanus.
Sites.
Irvingtonian/Early Rancholabrean: Archer (Jefferson 1991b).
Early/Early-Mid Wisconsin: Lost Valley (Harris 1993c).
Mid Wisconsin: Pendejo Cave (Harris 2003).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Blackwater Loc. No. 1 (Morgan and Lucas 2005); Charlies Parlor (Harris 1993c: cf.); Human Corridor (Harris 1993c: cf.); U-Bar Cave (Harris 1993c: cf.).
Late Wisconsin/Holocene: Burnet Cave (Schultz and Howard 1935; Fowlkes Cave (Dalquest and Stangl 1984b); Howell's Ridge Cave (Harris 1993c); Pendejo Cave (Harris 2003); Sheep Camp Shelter (Harris 1993c).
Late Wisconsin/Holocene: Isleta Cave No. 1 (Harris 1993c) Rejected
Literature. Dalquest and Stangl 1984b; Harris 1993c, 2003; 1991b; Morgan and Lucas 2005; Schultz and Howard 1935.
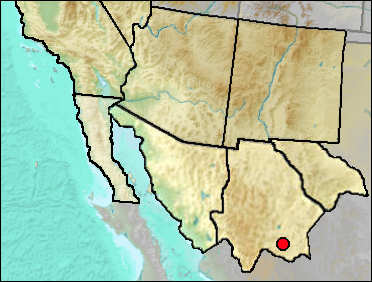
The region about Jimenez Cave is one of the few areas where Dipodomys nelsoni and D. spectabilis are sympatric (Best 1988). Both species are large with only modest size differences between the two.
Sites.
Mid/Late Wisconsin/Holocene: Jimenez Cave (Messing 1986).
Literature. Best 1988; Messing 1986.
Dipodomys panamintinus currently has a rather limited range, mostly in the Mojave Desert.
Sites.
Mid/Late Wisconsin: Pintwater Cave (Hockett 2000).
Late Wisconsin: Dove Springs Wash (Jefferson 1991b: cf.).
Literature. Hockett 2000.
The Banner-tailed Kangaroo Rat is enough larger than D. merriami and D. ordii as to usually be identifiable on size alone in the eastern part of the region. The current northern limits of distribution is near the Four-corners area of New Mexico.
This is primarily a grassland and desert-grassland animal that extends into desert. However, its widespread occurrence in late Wisconsin times suggests a wider ecological niche than is apparent today. It generally builds a large (6 ± ft diameter) mound of dirt with several burrow entrances.


Fig. 1. Dipodomys spectabilis. Fig. 2. Mound of Dipodomys spectabilis. Figs. 1 and 2 after Bailey (1931).
Sites.
Early/Early-Mid Wisconsin: Lost Valley (Harris 1993c).
Mid Wisconsin: Pendejo Cave (Harris 2003); Tank Trap Wash (Van Devender et al. 1987); U-Bar Cave (Harris 1987: cf.).
Mid/Late Wisconsin: Animal Fair (Harris 1993c); Dark Canyon Cave (Tebedge 1988); Navar Ranch (Van Devender et al. 1987).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Balcony Room (UTEP); Bison Chamber (Harris 1970a); Dust Cave (Harris and Hearst 2012); Human Corridor (Harris 1993c); Pendejo Cave (Harris 1993c); TT II (Harris 1993c); Pendejo Cave (Harris 2003); U-Bar Cave 13-14 ka (Harris 1989); cf.); U-Bar Cave 14-15 ka (Harris 1989); U-Bar Cave 15-18 ka (Harris 1989; cf.); U-Bar Cave 18-20 ka (Harris 1989: cf.).
Late Wisconsin/Holocene: Beyond Bison Chamber (Harris 1970a); Conkling Cavern (UTEP); Deadman Cave (Mead et al. 1984); Fowlkes Cave (Dalquest and Stangl 1984b); Howell's Ridge Cave (Harris 1993c); Isleta Cave No. 1 (UTEP); Pendejo Cave (Harris 2003).
Holocene:Isleta Cave No. 2 (Harris 1993c), reject as Holocene.
Literature. Bailey 1931; Dalquest and Stangl 1984b; Harris 1970a, 1987, 1989, 1993c, 2003; Harris and Hearst 2012; Mead et al. 1984; Tebedge 1988; Van Devender et al. 1987.
Last Update: 7 Jan 2016