Class Mammalia
Order Rodentia
Family Cricetidae
Subfamily Neotominae
Neotoma sp.—Woodrats // Neotoma (Paraneotoma) sp.—Paraneotoma Woodrats // Neotoma albigula—White-throated Woodrat // Neotoma cinerea—Bushy-tailed Woodrat // Neotoma devia/lepida—Arizona/Desert Woodrat // Neotoma findleyi—Findley's Woodrat // Neotoma floridana—Eastern Woodrat // Neotoma fossilis—Fossil Woodrat // Neotoma fuscipes—Dusky-footed Woodrat // Neotoma goldmani—Goldman's Woodrat // Neotoma lepida—Desert Woodrat // Neotoma leucodon—White-toothed Woodrat // Neotoma mexicana—Mexican Woodrat // Neotoma micropus—Southern Plains Woodrat // Neotoma pygmaea—Pygmy Woodrat // Neotoma pygmaea/stephensi—Pygmy or Stephen's Woodrat // Neotoma species c—Unnamed Woodrat c // Neotoma species e—Unnamed Woodrat e // Neotoma stephensi—Stephen's Woodrat // Neotoma taylori—Taylor's Woodrat
Woodrats, also known as packrats and as traderats, form a significant portion of Southwestern Pleistocene faunas. One reason for this is that most of the fossil faunas have been recovered from rock shelters and caves, prime habitat for most species of Neotoma.
Woodrats are relatively large rodents in the general size range of the Old World introductions, the Norway and Black rats (Rattus). Woodrats are more attractive to many people than Rattus, however, due likely to the relatively large eyes, haired tails, and often attractive pelage.
Eight species currently occur in the region (Harris 1984b): Neotoma albigula, N. cinerea, N. devia, N. lepida, N. leucodon, N. mexicana, N. micropus, and N. stephensi. Neotoma devia and N. lepida were recognized as separate species after most records were published and, since their ranges overlap, are given as N. devia/lepida, indicating that which species was present is unknown.
All are known as fossils from the region, although N. leucodon is only assumed on geographic grounds. Two other living species not currently known from the region also are recorded: N. floridana and N. goldmani, though the latter only tentatively (Harris 1984b), and two extinct species have been described from sites in New Mexico (Harris 1984d).
Neotoma leucodon has been recognized as a separate species only recently on the basis of genetic data. Previously, it has been included in N. albigula, and although there appears to be some average differences in size, there is no known character of teeth or skeleton that will allow identification of fossil material. The distribution as currently known lies east of the Rio Grande and Rio Conchos, whereas the distribution of N. albigula is west of those rivers except in a small area around El Paso, TX. Fossil "albigula-like" woodrats from the current range of N. leucodon have been assigned to that species, but it should be recognized that this is done on strictly geographic grounds.
Although generally quite easy to identify in the flesh, fossil remains have been a challenge. Post-cranial parts generally are identifiable only to genus. Maxillae and dentaries often survive with one or more teeth in place, and isolated teeth are common in most deposits. Several characteristics of teeth, especially m1, frequently will allow placement to species (Harris 1984b).
 Two
species groups can be recognized on the basis of presence or development of the
anterior lateral dentine tract on m1 (Harris 1984b). Among modern forms, N. cinerea,
N. mexicana, and N. stephensi have the tract well developed (Fig. 1),
whereas N. albigula, N. floridana, N. leucodon, N. devia, N. lepida, and N.
micropus generally do not. The extension of a smaller extension of dentine at the
base of the second loph of m1 also normally is better developed in the dentine-tract
group. Other teeth may show incipient tracts that are better developed than in the
other group.
Two
species groups can be recognized on the basis of presence or development of the
anterior lateral dentine tract on m1 (Harris 1984b). Among modern forms, N. cinerea,
N. mexicana, and N. stephensi have the tract well developed (Fig. 1),
whereas N. albigula, N. floridana, N. leucodon, N. devia, N. lepida, and N.
micropus generally do not. The extension of a smaller extension of dentine at the
base of the second loph of m1 also normally is better developed in the dentine-tract
group. Other teeth may show incipient tracts that are better developed than in the
other group.
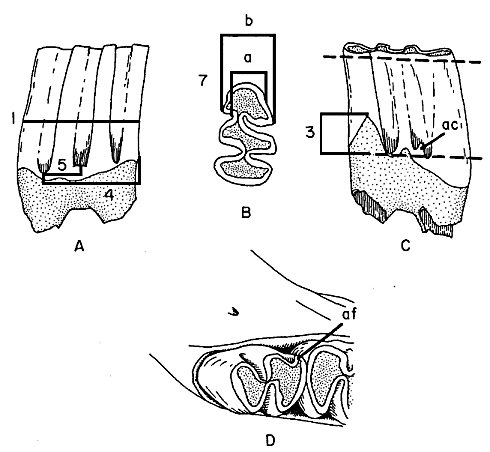
Fig. 1. Right m1 of fossil Neotoma cinerea showing a very well developed anterior lateral dentine tract (ivory color) and a typical extension dorsally of dentine on the second loph. Fossilization of this specimen has turned the enamel nearly black.
Harris (1984b) did an intensive survey of woodrats and included a number of ways to discriminate among the species. The lower first molar is the most useful tooth element for identification. Figure 2 shows measurements on m1 and some qualitative characters. Measurements consistently taken on lower jaw elements: length of alveolar cheek-tooth row (LG-ALV); mid-length of m1 (LG-M 1); greatest width of loph 2 of ml (WD-MI); height of antero-lateral dentine tract of ml (TRACT); (distance from base of lingual fold 2 to anterior face of ml (ANT-F2); distance from base of lingual fold 1 to base of fold 2 of ml (Fl-F2); development of the antero-internal reentrant fold of loph 1, m1 (FOLD); and (a ratio comparing depth of the ml antero-internal reentrant fold to the width of ml (RATIO) (Harris 1984b).
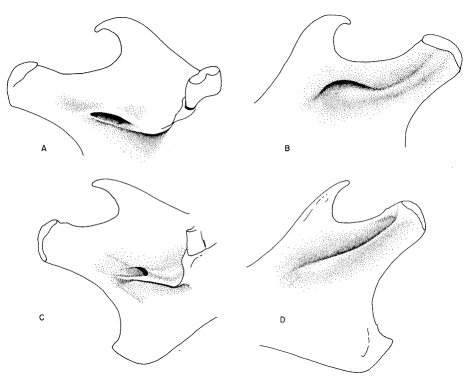
Fig. 2. A) Lingual view of left ml of Neotoma cinerea, showing method of taking LG-M1, ANT-F2, and F1-F2. B) Occlusal view of left ml of Neotoma albigula, showing measurements for calculation of RATIO (a/b). C) Labial view of left ml of N. cinerea, showing method of measuring the dentine tract (TRACT) and presence of accessory cusp (ac). D) Occlusal view of left m3 of Neotoma goldmani, showing accessory fold (af). After Harris (1984b).
 Qualitative
features useful in identification include the development of the incisive capsule and
the mandibular foramen. Figure 3 shows typical conditions. A and B represent the
mandibular foramen and incisive capsule in N. albigula and C and D represent the
same in N. goldmani. In general, the features seen in A and B are found in the
non-dentine tract species and those in C and D in the dentine-tract group. There is,
however, some overlap.
Qualitative
features useful in identification include the development of the incisive capsule and
the mandibular foramen. Figure 3 shows typical conditions. A and B represent the
mandibular foramen and incisive capsule in N. albigula and C and D represent the
same in N. goldmani. In general, the features seen in A and B are found in the
non-dentine tract species and those in C and D in the dentine-tract group. There is,
however, some overlap.
Fig. 3. A and B show the generally somewhat down-oriented mandibular foramen and the prominent incisive capsule in members of the non-dentine group. C and D show the more medially directed mandibular foramen and the less prominent incisive capsule seen in members of the dentine tract group. After Harris (1984b).
For a scatter diagram showing size differences of m1 among species with a low anterior lateral dentine tract, see the N. micropus account.
Literature. Harris 1984b; Harris 1984d.
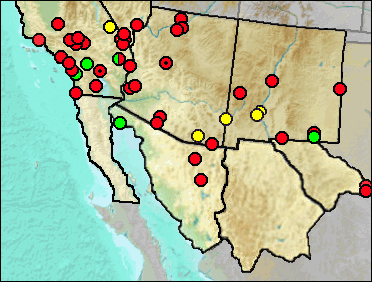
Teeth, and some skeletal elements, often can be told to genus, but not to species. This is the case with remains from the sites listed below.
Sites.
Blancan: Silver Canyon (Morgan et al. 2009)
Late Blancan: Caballo (Morgan et al. 2011); Curtis Ranch (Lindsay 1984); Kelly Canyon (Morgan et al. 2011); Tecopa Lake Beds (Woodburne and Whistler 1991); Virden (Morgan et al. 2008).
Irvingtonian: El Golfo (Croxen et al. 2007 [Neotoma (Neotoma) and Neotoma (Hodomys)]; Elsinore: Microtus/Mammuthus (Pajak et al. 1996).
Early Irvingtonian: El Casco, San Timoteo Badlands (Albright 2000).
Medial Irvingtonian: Slaughter Canyon Cave (Morgan and Harris 2015).
Late Irvingtonian: Elsinore: Pauba Formation (Pajak et al. 1996).
Irvingtonian/Rancholabrean: Cadiz (Jefferson 2014).
Rancholabrean: Bedford Properties (Jefferson 1991b); Big Rock (Mead et al. 2005); Bow Willow (Jefferson 2014); Centennial Parkway, Las Vegas Valley (Jefferson et al. 2015); Century City, Los Angles (Jefferson 1991b); Clay Mine Road (Jefferson 2014); Detention Basin, Upper Las Vegas Wash (Jefferson et al. 2015); Eagle Crest (Jefferson 2014); Edwards Air Force Base (Jefferson 2014); Helendale (Jefferson 1991b); Hoffman Road (Jefferson 2014); Park La Brea A (Jefferson 2014); Piute Valley (Jefferson 1991b); Morena Blvd. (Jefferson 1991b); Wilshire Blvd., 10580 The Wilshire, Los Angeles (Jefferson 1991b).
Sangamon: La Brisca (Van Devender et al. 1985); Naval Fuel Reserve Quarry (Jefferson 1991b).
Wisconsin: Arroyo de las Tinajas 1 (UTEP); Carpinteria (Wilson 1933); CC:5:1 (Mead et al. 2003); CC:5:5 (Mead et al. 2003); Costeau Pit (Jefferson 1991b); Glen Abbey, Bonita (Jefferson 1991b).
Mid Wisconsin: CC:5:2 (Mead et al. 2003); CC:5:3 (Mead et al. 2003); Devil Peak (Jefferson et al. 2015); Pacific City (Wake and Roeder 20009); Plumosa Mountains (Mead et al. 2005); Térapa (Mead et al. 2007).
Mid/Late Wisconsin: Rampart Cave (Lindsay and Tessman 1974); Sandblast Cave (Emslie 1988).
Late Wisconsin: Antelope Cave (Jefferson 1991b); Bat Cave (Scarbrough 1986); Bida Cave (Mead et al. 2005); Blackwater Draw Fauna (Slaughter 1975); Bonfire Shelter (Frank 1968); Brass Cap Point (Mead et al. 1983); Corn Creek Packrat Midden (Jefferson et al. 2015); Cueva Quebrada (Lundelius 1984); Falling Arches (Jefferson 1991b); Flaherty Mesa #1 (Jefferson et al. 2015); Hance Canyon (Mead et al. 2005); Hueco Mountains (Van Devender et al. 1987); Montezuma's Head (Mead et al. 2005); Murray Springs (Hemmings 2007a); NAUQSP Locality 991 (Mead et al. 2003); New Water Mountains (Mead et al. 1983); Park Place, Irvine (Jefferson 2014); Red Tail Peak Midden (Jefferson 1991b); Shinumo Creek (Van Devender et al. 1977); Skull Cave (Emslie 1988); Spires SP2, Sheep Range (Jefferson et al. 2015); Stanton's Cave (Olsen and Olsen 1984); Valley Wells (Springer et al. 2010); Ventana Cave (Lance 1959); Vulture Cave (Mead and Phillips 1981).
Rancholabrean/Holocene: Arvin Landfill (Jefferson 2014).
Late Wisconsin/Holocene: Luz Foundation (Jefferson 1991b); Schuiling Cave (Jefferson 1991b).
Literature. Albright 2000; Croxen et al. 2007; Emslie 1988; Frank 1968; Harris 1993c; Hemmings 2007a; Jefferson 1991b, 2014; Jefferson et al. 2015; Lance 1959; Lindsay 1984; Lindsay and Tessman 1974; Lundelius 1984; Mead et al. 1983, 2003, 2005, 2007; Mead and Phillips 1981; Morgan 2003; Morgan et al. 2008; Morgan et al. 2009; Morgan et al. 2011; Morgan and Harris 2015; Olsen and Olsen 1984; Pajak et al. 1996; Reynolds, Reynolds, and Bell 1991; Scarbrough 1986; Slaughter 1975; Springer et al. 2010; Van Devender et al. 1977, 1987, 1985; Wake and Roeder 2009; Wilson 1933; Woodburne and Whistler 1991.
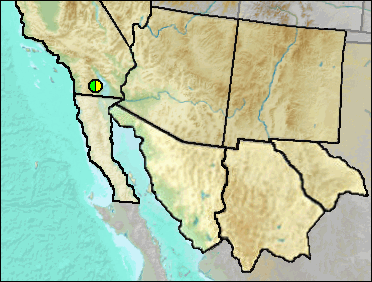
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Murray 2008.
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Murray 2008.
Last Update: 3 Feb 2016