Class Mammalia
Order Rodentia
Family Heteromyidae
Chaetodipus/Perognathus—Spiny or Silky Pocket Mice // Perognathus sp—Silky Pocket Mice // Perognathus flavus—Silky Pocket Mouse // Perognathus gidleyi—Gidley's Pocket Mouse // Perognathus inornatus—San Joaquin Pocket Mouse // Perognathus longimembris—Little Pocket Mouse // Perognathus merriami—Merriam's Pocket Mouse
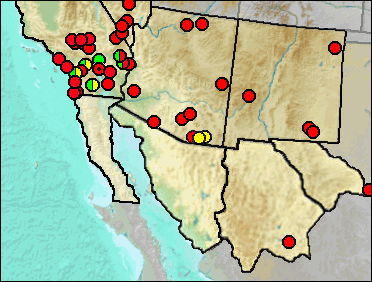
It's not clear whether some of the larger silky pocket mice can be separated from smaller species of Chaetodipus on the basis of the types of material usually available as fossils, let alone into morphological species of Perognathus. At the time many of the records below were published, all of our pocket mice were considered to belong to Perognathus—thus there is no way to know for these which genus was represented without examination of the specimens involved, and quite likely not then.
Sites.
Late Blancan: California Wash (Morgan and White 2005); Curtis Ranch (Lindsay 1984); Jack Rabbit Trail, San Timoteo Badlands (Albright 2000).
Late Blancan/Early Irvingtonian: Elsinore: Mimomys (Pajak et al. 1996).
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Irvingtonian: El Casco, San Timoteo Badlands (Albright 2000); Elsinore: Microtus/Mammuthus (Pajak et al. 1996).
Early Irvingtonian: Gypsum Ridge (Wagner and Prothero 2001).
Irvingtonian/Early Rancholabrean: Archer (Jefferson 1991b).
Late Irvingtonian: Elsinore: Pauba Formation (Pajak et al. 1996).
Irvingtonian/Rancholabrean: Cadiz (Jefferson 2014).
Rancholabrean: Bedford Properties (Jefferson 1991b); Bitter Springs Playa (Jefferson 2014: Cf.); Centennial Parkway, Las Vegas Valley (Jefferson et al. 2015); Clay Mine Road (Jefferson 2014); Clay Mine Road (Jefferson 2014: small); Cool Water Coal Gasification Solid Waste Site (Jefferson 1991b); Desert Sunlight Project (Jefferson 2014); Detention Basin, Upper Las Vegas Wash (Jefferson et al. 2015: large and small species); Glen Abbey, Bonita (Jefferson 1991b); Helendale (Jefferson 1991b); Hoffman Road (Jefferson 2014); Mescal Cave (Jefferson 1991b); Morena Blvd. (Jefferson 1991b); National City West (Jefferson 2014); Siphon 11 (Jefferson 1991b); Tule Springs (Springer et al. 2005); Tramperos Creek (Morgan and Lucas 2005).
Rancholabrean/Early Holocene: Metro Rail Universal City Station (Jefferson 2014).
Late Pleistocene: Wanis View (Jefferson 2014).
Early/Early-Mid Wisconsin: Lost Valley (Harris 1993c).
Mid Wisconsin: Devil Peak (Jefferson et al. 2015); Papago Springs Cave (Czaplewski and Mead et al. 1999); Plumosa Mountains (Mead et al. 1983); Screaming Neotoma Cave (Glennon 1994); Shonto (Mead et al. 2005).
Mid/Late Wisconsin: Dark Canyon Cave (Harris 1993c); Diamond Valley (Springer et al. 2009); Pintwater Cave (Jefferson et al. 2015); Sandblast Cave (Emslie 1988); Tsuma Properties, San Clemente (Jefferson 2014).
Mid/Late Wisconsin/Holocene: Jimenez Cave (Messing 1986).
Late Wisconsin: Alamo Canyon (Mead et al. 1983); Bonfire Shelter (Frank 1968); Cueva Quebrada (Lundelius 1984: possibly both genera); Falling Arches (Jefferson 1991b); Murray Springs (Mead et al. 2005); Picacho Peak (Van Devender et al. 1991); Potosi Mountain (Mead and Murray 1991).
Late Wisconsin/Holocene: Bat Cave (Scarbrough 1986); Calico Lakes (Jefferson 1991b); Wolcott Peak (Mead et al. 1983); Luz Foundation (Jefferson 1991b); Luz Solar Trough (Jefferson 1991b); Schuiling Cave (Jefferson 1991b; Solar One (Jefferson 1991b); Whipple Mountains Midden (Jefferson 1991b).
Literature. Albright 2000; Czaplewski and Mead et al. 1999; Emslie 1988; Frank 1968; Glennon 1994; Harris 1993c; 1991b, 2014; Jefferson et al. 2015; Lindsay 1984; Mead et al. 1983; Mead et al. 2005; Lundelius 1984; Mead and Murray 1991; Messing 1986; Morgan and Lucas 2005; Pajak et al. 1996; Reynolds, Reynolds, and Bell 1991; Scarbrough 1986; Springer et al. 2005, 2009; Van Devender et al. 1991; Wagner and Prothero 2001.
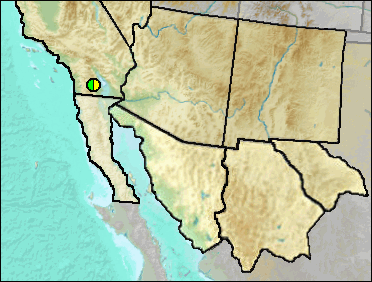
According to Murray (2008), a diagnosis was included in a M.S. thesis by Gensler (2002), but the taxon has not been formally named nor published.
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Gensler 2002; Murray 2008.
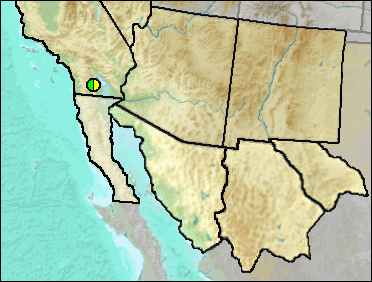
According to Murray (2008), a diagnosis was included in a M.S. thesis by Gensler (2002), but the taxon has not been formally named nor published.
Sites.
Late Blancan/Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Gensler 2002; Murray 2008.
The genus Perognathus was long divided into two subgenera: P. (Chaetodipus) and P. (Perognathus) until they were deemed to be enough different as to represent two separate genera (Hafner and Hafner 1983).
Perognathus shares many characteristics with Chaetodipus, but have auditory bullae even more enlarged and have fine hairs making up the pelage rather than the relatively coarse hairs characteristic of the Spiny Pocket Mice. Our species also are small, with Perognathus flavus being the smallest North American rodent.
Dalquest and Stangl (1984b) made a number of measurements of pocket mice and concluded that most or all of dentaries with the alveolar cheek tooth row less than 3.15 mm likely represented P. flavus.
Frey (2004) recognized four species of silky pocket mice in New Mexico, whereas apparently Dalquest and Stangl (1984b) only considered one possibility. The problem with silky pocket mice in our region is that species have been split and merged over the fairly recent past. The following situation is not necessarily agreed upon by all. Following Frey (2004), Perognathus merriami is recognized as separate from P. flavus on the basis of genetic, but not morphological, information. Perognathus apache has been considered a separate species, then as a subspecies of Perognathus flavescens, and now as separate once again. Perognathus apache/flavescens is slightly larger than P. flavus, and the latter apparently is indistinguishable from P. merriami on the basis of size or osteological character. Which of these species is represented by the records below is problematic; records are given as recorded in the literature except where otherwise stated.
Literature. Dalquest and Stangl 1984b; Frey 2004; Hafner and Hafner 1983.
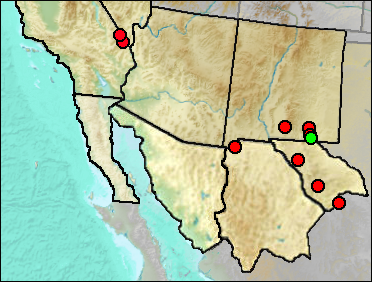
These records are of specimens originally identified as Perognathus (Perognathus), thus ruling out Chaetodipus (assuming correct identifications to subgenus).
Medial Irvingtonian: Slaughter Canyon Cave (Morgan and Harris 2015).
Early/Early-Mid Wisconsin: Rm Vanishing Floor (Harris 1993c).
Mid Wisconsin:Tunnel View #8A(2) (Van Devender and Bradley 1990).
Mid/Late Wisconsin: Animal Fair (Harris 1993c).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Antelope Cave (Reynolds, Reynolds, Bell, and Pitzer 1991); Bison Chamber (Harris 1970a); Charlies Parlor (Harris 1989); Pendejo Cave (Harris 2003); TT II (Harris 1993c); U-Bar Cave 13-14 (Harris 1989); U-Bar Cave 14-15 ka (Harris 1989); U-Bar Cave 15-18 ka (Harris 1989); U-Bar Cave 18-20 ka (Harris 1989).
Late Wisconsin/Holocene: Beyond Bison Chamber (Harris 1970a); Entrance Chamber (UTEP); Kokoweef Cave (Reynolds, Reynolds, et al. 1991); Pendejo Cave (Harris 2003).
Literature. Harris 1970a, 1989, 1993c, 2003; Jefferson 1991b; Morgan and Harris 2015; Reynolds, Reynolds, et al. 1991; Reynolds, Reynolds, Bell, and Pitzer 1991; Van Devender and Bradley 1990.
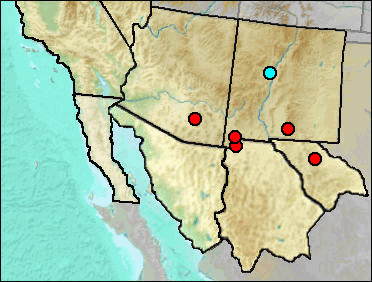
El Paso is the type locality of P. flavus.
The following are sites where P. flavus is identified in the literature. As noted in the generic account, however, at least the possibility of other species must be acknowledged.
Sites.
Mid Wisconsin: Pendejo Cave (UTEP: cf.); U-Bar Cave (Harris 1987: cf.).
Late Wisconsin/Holocene: Deadman Cave (Mead et al. 1984; cf.); Fowlkes Cave (Dalquest and Stangl 1984b); Howell's Ridge Cave (Harris 1993c); Pendejo Cave (UTEP: cf.).
Holocene Isleta Cave No. 1 (Harris 1993c: cf.) and Isleta Cave No. 2 (Harris 1993c) rejected as not a Pleistocene occurrence.
Literature. Dalquest and Stangl 1984b; Harris 1987, 1993c; Mead et al. 1984.
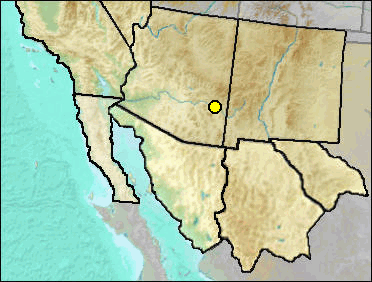
This is a pocket mouse the size of the extant P. formosus, originally described from the Rexroad fauna of Meade County, Kansas (Hibbard 1941).
Sites.
Late Blancan: 111 Ranch (Morgan and White 2005).
Literature. Hibbard 1941; Morgan and White 2005.
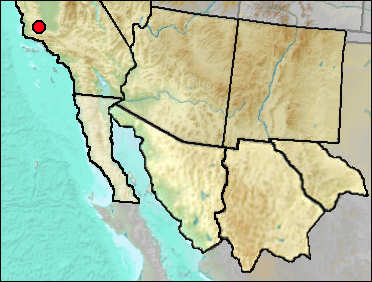
Current geographic distribution is limited to central California.
Sites.
Mid Wisconsin: McKittrick (Schultz 1937: cf.).
Literature. Schultz 1937.
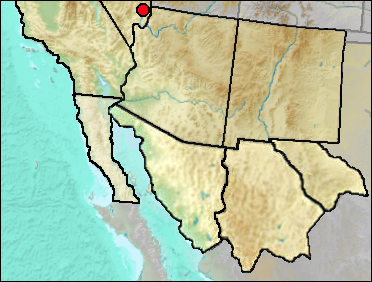
The current distribution of this small pocket mouse is Great Basin south into Mexico.
Sites.
Rancholabrean: Clay Mine Road (Jefferson 2014).
Mid Wisconsin: Glendale (Jefferson et al. 2015).
Literature. Jefferson et al. 2015
Perognathus merriami has at times been considered as conspecific with P. flavus and at times as a separate species. Currently it is recognized as separate, mostly on the basis of biochemical genetic data. However, Tebedge (1988), citing Lundelius, noted that "P. merriami collected from Pratt Cave differed from P. flavus in the greater posterior turning of the anterolabial cusps of M1 and M2, the lesser anteroposterior compression of the cross lophs of M1 and M2, the smaller size of the posterolabial cusps of M1 and M2 (especially M3), and the relatively deeper mandible. P. merriami differs from P. flavescens in the larger P4 and the presence of an external cingulum joining the two lophs on M1 and M2 (Lundelius, 1979)."
Sites.
Mid/Late Wisconsin: Dark Canyon Cave (Tebedge 1988).
Late Wisconsin: Muskox Cave (Logan 1981; Harris 1993c as P. flavus).
Literature. Logan 1981; Tebedge 1988.
Last Update: 28 May 2015