Class Mammalia
Order Rodentia
Family Cricetidae
Subfamily Neotominae
Peromyscus sp.—Deer Mice // Peromyscus anyapahensis—Anacapa Island Mouse // Peromyscus boylii—Brush Mouse // Peromyscus californicus—California Mouse // Peromyscus complexus—Complex Mouse // Peromyscus crinitus—Canyon Mouse // Peromyscus eremicus—Cactus Mouse // Peromyscus hagermanensis—Hagerman Mouse // Peromyscus imperfectus—Imperfect Mouse // Peromyscus leucopus—White-footed Mouse // Peromyscus maniculatus—Deer Mouse // Peromyscus maximus—Maximum-sized Mouse // Peromyscus nasutus—Northern Rock Mouse // Peromyscus nesodytes—Giant Deer Mouse // Peromyscus laceianus—Northern White-ankled Mouse // Peromyscus truei—Piñon Mouse
Many areas in our region today have four or more species of Peromyscus. Unfortunately, identification to species may be difficult with an intact animal in hand; difficulties ramify with fragmentary osteological or dental material. Thus many records are given only as Peromyscus or even as "peromyscine" where material may not allow discrimination between Onychomys, Reithrodontomys, and Peromyscus.
Dalquest and Stangl (1983) did an extensive study of seven species of Peromyscus that were deemed as possible members of early Holocene and Pleistocene faunas in Trans-Pecos Texas. As they pointed out, dentary rami and first lower molars are most often preserved, and thus they concentrated on those elements for discriminatory character states. They found the nature of the incisor-base capsule, the length of m1, alveolar length of the molar row, and details of the enamel pattern of m1 to be the traits of major value; they also found that overlap between species make it difficult to identify a single lower jaw to species in many cases. Nevertheless (p. 6):
In most features, however, the overlap between some species was slight, resulting from one or two variant individuals. Given a large series of lower jaws of mixed species of Peromyscus from Trans-Pecos Texas, use of combinations of characters often made it possible to determine whether or not a species was present, to identify some individual jaws to species, and to narrow the identification of other jaws to one of two species.
In July and August of 2011, I concentrated on the identification of fossil Peromyscus, using Dalquest and Stangl's (1983) methods as a starting point. Alveolar length of the molar tooth row was taken according to their instructions (p. 4):
Because some teeth often are lost in fossil Peromyscus jaws, we depended on the alveolar length of the tooth row. Measurements were taken on the lingual side, with the jaw slightly tilted, so that the margins of the alveoli were visible.
Likewise, their coding for the anteroconid of m1 was utilized (p. 4):
Typically, the anteroconid of ml of Peromyscus bears a slight notch or groove on the anterdorsal [sic!] surface of the unworn tooth. This notch is often absent and even when present, may be so shallow that it is worn away in early stages of attrition; this we consider Grade O. In some species the notch, though narrow, is persistent and moderately deep until rather late stages of moderate tooth wear; this is Grade 1. In Grade 1, the groove is so narrow that there is little or no space between the enamel surface. In Grade 2 there is a distinct valley separating the anteroconid into inner and outer parts. This valley, with the valley between anteroconid and metaconid, may give the trigonoid a trefoil or clover-leaf appearance.
Length of m1 was taken in dorsal view from the most anterior portion of the tooth to the medial meeting with m2. The
incisor capsule was coded from 0 to 3: 0, a smooth curve seen from above; 1, a very
slight bulge both from above and in lateral view; 2, a large bulge dorsally, but
blending into the ramus posteriorly; and 3, a large bulge (projection) as in
"2", but also separated postero-dorsally from the adjacent wall by an
indentation (Fig. 1). 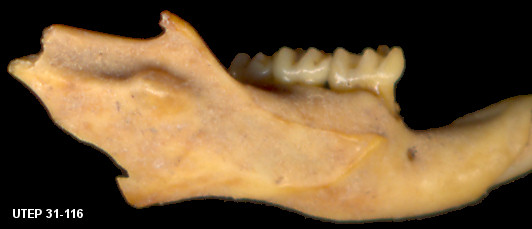
A fifth measurement was added: distance from the posterior alveolus of m3 to the anterior opening of the mandibular foramen, with both in approximately the same plane. Linear measurements were taken with an eyepiece micrometer. All linear measurements were rounded to the nearest 0.5 mm except recorded as 0.25 mm when there was no clear rounding decision; a similar strategy was used for the judgmental categories (capsule and anteroconid). Precision of measurements was relatively low due to issues of orientation and wear.
Fig. 1. Dentary of Peromyscus maniculatus showing the incisor capsule coded as "3". This typical form is shown by UTEP 31-116, Dry Cave.
Samples of modern specimens were measured and fossil specimens compared to modern samples by discriminant analysis, at times supplemented by cluster and principal component analyses, using PAST software (Hammer et al. 2001). Limitations include the lack of modern specimens of P. gratus (Osgood's Mouse) from western New Mexico, formerly included in P. truei, and a limited sample of regional modern P. crinitus (supplemented, however, with specimens from Nevada and California). Osgood's Mouse likely is of concern only in sites west of the Rio Grande.
The new identifications given here are considered well founded at the level given unless commented upon otherwise. However, identifications to species in the literature should be taken with a large grain of salt unless individually defended appropriately. Some literature records are commented upon in the species accounts. Some early, overly-optimistic identifications by myself are rejected below (blue dots).
As noted by Dalquest and Stangl (1983), the Peromyscus of our region tend to fall into two groups: those with an enlarged incisor-base capsule and those without appreciable enlargement. Peromyscus crinitus, P. eremicus, P. leucopus, and P. maniculatus fall into the first group, whereas P. boylii, P. nasutus, P. pectoralis, and P. truei fall into the second. However, occasional individuals are atypical; among the species that tend to have enlarged capsules, capsule development in the sample runs from 0 to 2 (P. crinitus), 1 to 3 (P. leucopus), and 0 to 3 (P. eremicus, P. maniculatus). On the other hand, capsule development only runs from 0 to 1 in the sample of the other species. There is some indication that capsule development in the first group may be partly dependent on age, young animals having lesser development.
Literature. Dalquest and Stangl 1983; Hammer et al. 2001.
Identification of deer mice to only the generic level is of little value (one species or another occurs almost everywhere), but site records are given here for the sake of completeness.
Sites.
Irvingtonian: Anza-Borrego (Murray 2008); Elsinore: Microtus/Mammuthus (Pajak et al. 1996).
Late Irvingtonian: Elsinore: Pauba Formation (Pajak et al. 1996).
Rancholabrean/?Early Holocene: Mitchell Caverns (Jefferson 1991b); Metro Rail Universal City Station (Jefferson 2014).
Mid/Late Wisconsin/Holocene: Jimenez Cave (Messing 1986).
Literature. Czaplewski and Mead et al. 1999; Czaplewski and Peachey 2003; Emslie 1988; Glennon 1994; Harris 1989, 1993c, 2003; Harris and Hearst 2012; Hockett 2000; Jefferson 1991b, 2014; Jefferson et al. 2015; Lindsay 1984; Lindsay and Tessman 1974; Lundelius 1984; Logan 1981; Lundelius 1984; McDonald et al. 2004; Mead and Murray 1991; Mead and Phillips 1981; Mead et al. 1983; Mead et al. 1984; Mead et al. 2003; Mead et al. 2005; Messing 1986; Morgan and Lucas 2005; Murray 2008; Olsen and Olsen 1984; Pajak et al. 1996; Reynolds, Reynolds, and Bell 1991; Reynolds, Reynolds, et al. 1991; Reynolds, Reynolds, Bell, and Pitzer 1991; Rogers et al. 2000; Scarbrough 1986; Springer et al. 2010; Stearns 1942; Van Devender and Bradley 1990; Van Devender et al. 1977a, 1987, 1989; Woodburne and Whistler 1991.
Last Update: 27 Jun 2015