Class Mammalia
Order Carnivora
Family Procyonidae
Bassariscus sp.—Ringtails // Bassariscus astutus—Ringtail // Bassariscus casei—Case's Ringtail // Procyon sp.—Raccoons // Procyon lotor—Raccoon // Procyon rexroadensis—Rexroad Raccoon
Procyonids are medium to medium-large mammals generally considered to be omnivores, though the Ringtail veers toward the carnivorous side.
Three species of procyonids occur in our regions: Bassariscus astutus (Ringtail), Nasua narica (White-nosed Coati), and Procyon lotor (Raccoon). The coati barely enters into our region today in southwestern New Mexico and southeastern Arizona. The Raccoon, though quite widespread today in our region, is scantily represented as a fossil. In contrast, Ringtails are common in both the modern-day region and as fossils.
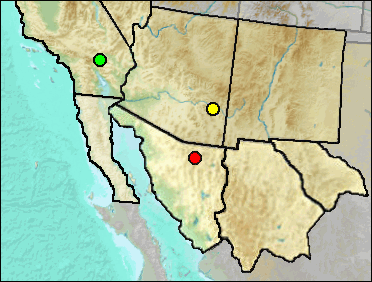
Sites.
Late Blancan: 111 Ranch (Morgan and White 2005: ?).
Early Irvingtonian: Gypsum Ridge (Wagner and Prothero 2001).
Sangamon: La Brisca (Van Devender et al. 1985).
Literature.
Morgan and White 2005; Van Devender et al. 1985; Wagner and Prothero 2001.
Synonyms. Bassariscus sonoitensis Skinner 1942.
 Ringtails are common inhabitants of the
Southwestern roughlands, from low desert to montane forests. They also are fairly
common members of late Pleistocene faunas. These agile members of the Raccoon family
tend to be somewhat more carnivorous than Raccoons. They are at home wherever there are
climbable surfaces, easily scaling near-vertical rock faces. They are known to
occasionally venture beyond the twilight zone of caves.
Ringtails are common inhabitants of the
Southwestern roughlands, from low desert to montane forests. They also are fairly
common members of late Pleistocene faunas. These agile members of the Raccoon family
tend to be somewhat more carnivorous than Raccoons. They are at home wherever there are
climbable surfaces, easily scaling near-vertical rock faces. They are known to
occasionally venture beyond the twilight zone of caves.
Fig. 1. Ringtail. Photograph courtesy of the U.S. Fish & Wildlife Service.
Bassariscus sonoitensis was described from Papago Springs cave, Santa Cruz County, Arizona (Skinner 1942). Czaplewski et al. (1989) used radiometric dating to determine that the age of the deposits predated the late Wisconsin, thus suggesting that perhaps B. sonoitensis was ancestral to B. astutus. Harris (1990b) examined the relationship between B. sonoitensis and B. astutus, arriving at the conclusion that the former should be considered a synonym of B. astutus, although possibly deserving of temporal subspecific status based on it being characteristically within or near the upper limits of the present populations on the characters of tooth length and width and of robustness of muzzle.
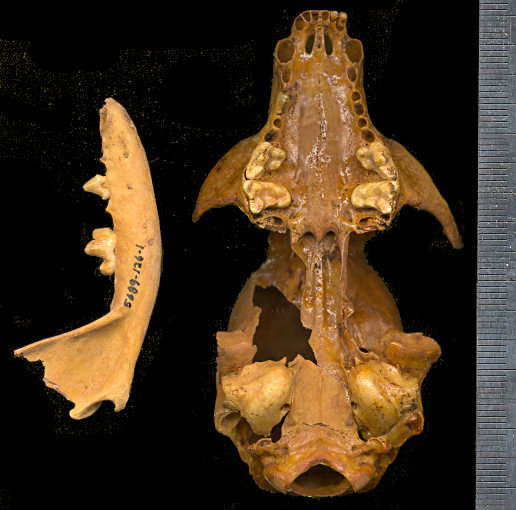 Fig. 2. Ventral view of the
skull and lateral view of the dentary of Bassariscus astutus from U-Bar Cave
(UTEP 5689.140.53). The skull originally was identified as Bassariscus sonoitensis
and formed the basis for further research concerning the relationship between B.
astutus and B. sonoitensis. Scale in mm.
Fig. 2. Ventral view of the
skull and lateral view of the dentary of Bassariscus astutus from U-Bar Cave
(UTEP 5689.140.53). The skull originally was identified as Bassariscus sonoitensis
and formed the basis for further research concerning the relationship between B.
astutus and B. sonoitensis. Scale in mm.
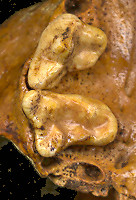 Fig. 3. Enlargement of right P4
and M1 of Bassariscus astutus, same individual as in Fig. 2.
Fig. 3. Enlargement of right P4
and M1 of Bassariscus astutus, same individual as in Fig. 2.
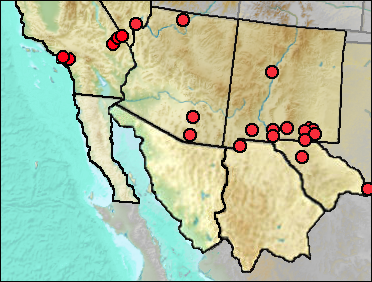
Mid and late Wisconsin recorded only as Bassariscus have been entered here as B. astutus.
Sites.
?Late Irvingtonian/Rancholabrean: Emery Borrow Pit (Jefferson 1991b: cf.).
Rancholabrean: Anthony Gap Cave (UTEP); Mescal Cave (Jefferson 1991b); Papago Springs Cave (Skinner 1942); Spotted Ridge (Jefferson et al. 2015).
Rancholabrean/?Early Holocene: Mitchell Caverns (Jefferson 1991b).
Wisconsin: Big Manhole Cave (Harris 1993c).
Early/Early-Mid Wisconsin: Sabertooth Camel Maze (Harris 1993c).
Mid Wisconsin: Pendejo Cave (UTEP: cf. gen.); U-Bar Cave (Harris 1990b).
Mid/Late Wisconsin: Dark Canyon Cave (Tebedge 1988); Rampart Cave (Lindsay and Tessman 1974); Rancho La Brea (Akersten et al. 1979).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Animal Fair 18-20 ka (Harris 1989: Bassariscus sp.); Cueva Quebrada (Lundelius 1984: cf.); Harris' Pocket (Harris 1989: Bassariscus sp.); Muskox Cave (Logan 1981); Pendejo Cave (Harris 2003); U-Bar Cave 14-15 ka (Harris 1989: Bassariscus sp.); Upper Sloth Cave (Logan and Black 1979); Vulture Cave (Mead and Phillips 1981).
Late Wisconsin/Holocene: Baldy Peak Cave (Harris 1993c); Burnet Cave (Schultz and Howard 1935); Conkling Cavern (Harris 1993c); Deadman Cave (Mead et al. 1984); Kokoweef Cave (Reynolds, Reynolds, et al. 1991); Pendejo Cave (Harris 2003); Stanton's Cave (Olsen and Olsen 1984).
Literature.
Akersten et al. 1979; Czaplewski et al. 1989; Harris 1989, 1990b, 1993c, 2003; Jefferson et al. 2015; Lindsay and Tessman 1974; Logan 1981; Logan and Black 1979; Lundelius 1984; Mead and Phillips 1981; Mead et al. 1984; Olsen and Olsen 1984; Reynolds, Reynolds, et al. 1991; Schultz and Howard 1935; Skinner 1942; Tebedge 1988.
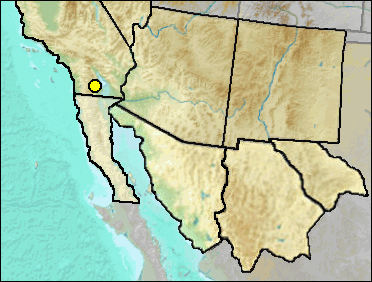
Hibbard (1952:379) described Bassariscus casei from the Rexroad fauna of Kansas, with the holotype consisting of a right ramus and part of the maxillary:
Diagnosis.-A Bassariscus the size of B. astutus (Lichtenstein) with the paraconid and protoconid, also the protoconid and metaconid, separated by wider valleys between these cusps than in the Recent species. The hypoconid and entoconid are not as well developed as in B. astutus. The paraconid of M2 is better developed and more centrally located on the anterior face of M2 than in B. astutus. P4 is distinct in that a well-developed cingulum extends from the hypocone to the base of the metacone, producing a distinct shelf on the inner side of the tooth, a condition not present in B. astutus although a less prominent shelf is present in Bassariscus sumichrasti (Saussure).
Sites.
Late Blancan: Anza-Borrego (Murray 2008).
Literature. Hibbard 1952; Murray 2008.
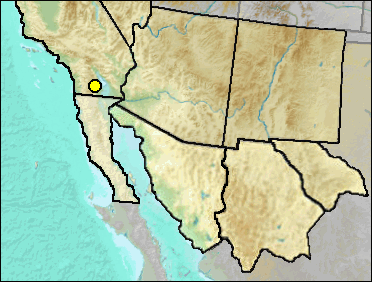
Sites.
Late Blancan: Anza-Borrego (Murray 2008).
Literature. Murray 2008.
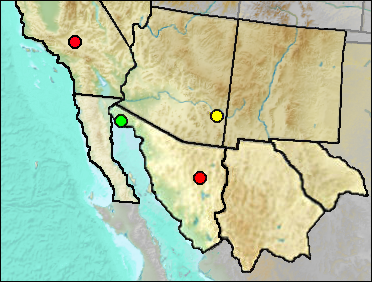
Sites.
Late Blancan: San Simon Fauna (Morgan and White 2005).
Irvingtonian: El Golfo (Croxen et al. 2007: gen. cf.).
Mid Wisconsin: Térapa (Mead et al. 2006).
Late Wisconsin/Holocene: Schuiling Cave (Jefferson 1991b).
Literature. Croxen et al. 2007; Jefferson 1991b; Morgan and White 2005.
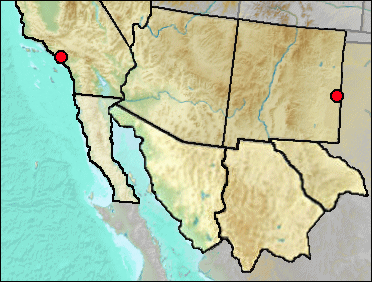
 Despite numerous records of fossil
Raccoons east of our region and present as fossils on the West Coast, and despite the presence of Raccoons in the central portions of our
region today, they are absent as fossils from the central region. Our only record on the east is from
Blackwater Draw, where several taxa otherwise known from east of our region show up in
the fossil record. The most likely reason for this pattern is eastern forms moving up
the gallery forests typical of the drainageways.
Despite numerous records of fossil
Raccoons east of our region and present as fossils on the West Coast, and despite the presence of Raccoons in the central portions of our
region today, they are absent as fossils from the central region. Our only record on the east is from
Blackwater Draw, where several taxa otherwise known from east of our region show up in
the fossil record. The most likely reason for this pattern is eastern forms moving up
the gallery forests typical of the drainageways.
The evidence strongly suggests recent invasion for most of the central region, probably as a result of human occupation of the area with the resultant food supply in the way of crops and garbage. Current distribution seems to be concentrated in river valleys and mountain forests in the vicinity of towns and campgrounds.
Fig. 1. Raccoon. Photograph courtesy of U.S. National Park Service.
Sites.
Late Wisconsin: Blackwater Draw Fauna (Slaughter 1975); Rancho La Brea (Akersten et al. 1979).
Literature.
Slaughter 1975; Akersten et al. 1979).
Kurtén and Anderson (1980) considered the Rexroad Raccoon to be the probable ancestor to P. lotor. They noted that the dental pattern is similar to that of P. lotor, but with the premolar-molar series narrower; the lower jaw is larger than that of the modern Raccoon.
Sites.
Late Blancan: Anza-Borrego (Murray 2008: cf.).
Literature. Kurtén and Anderson 1980; Murray 2008.
Last Update: 7 Apr 2015