Class Mammalia
Order Artiodactyla
Suborder Ruminantia
Family Antilocapridae
Croxen et al. (2007) listed Tetrameryx (Stockoceros) sp. and "gen. et sp. nov. † (4-horned pronghorn)" from El Golfo. Lacking further data, these are placed here as Stockoceros sp.
Sites.
Irvingtonian: Anza Borrego (Murray 2006: cf. gen.); El Golfo (Croxen et al. 2007).
Literature.
Croxen et al. 2007Murray 2006.
Synonyms. Tetrameryx conklingi, Tetrameryx onusrosagris, Stockoceros onusrosagris.
Stock (1930) originally described S. conklingi, from Shelter Cave, New Mexico, as Tetrameryx(?) conklingi. The type description is:
Specific characters.—Forked horn cores, arising from a common base, present on each side above and in part slightly behind orbit. Anterior horn core slightly stouter and longer than posterior; hinder not so greatly elongated as in Tetrameryx shuleri Lull. Base of anterior horn core relatively wider than in latter species. Horn cores divergent as in Tetrameryx, not parallel as in Capromeryx minor Taylor. Size distinctly smaller than T. shuleri and approximately two-thirds that of existing pronghorn. Metapodials relatively heavier than in Antilocapra americana (Ord).
Stock further characterized the species in 1932. Among new material from Shelter Cave, more of the holotype skull material was recovered. Measurements are given in Fig. 1 (Stock 1932).

Fig. 1. Facsimile of the table giving the measurements of the holotype of Stockoceros conklingi. After Stock (1932).
In 1934, Roosevelt and Burden described Tetrameryx onusrosagris from Papago Spring Cave, Arizona. The name Stockoceros was first applied by Frick (1937) as a subgenus of Tetrameryx, but most authors treat it as a genus (Kurtén and Anderson 1980).
Two species of Stockoceros have generally been recognized: S. conklingi and S. onusrosagris. However, there appears to be a question as to whether S. onusrosagris and S. conklingi represent one species or two. Roosevelt and Burden (1934:1), in their description of Stockoceros onusrosagris from Papago Springs Cave in southern Arizona, noted that "The horn cores, while notably larger and more branching, are in their general situation and form very much as those of Tetrameryx conklingi Stock of the Pleistocene of New Mexico", and they compared measurements of horn cores, teeth, and four limb bones with conklingi, with conklingi showing smaller measurements. Colbert and Chaffee (1939:17) compared Papago Springs Stockoceros extensively with Antilocapra, but limited their comments regarding it in comparison to S. conklingi to "it is much larger and the horn cores are more widely divergent." The Papago Springs Cave Stockoceros span a range of time from 246 ± 19 ka to 31.0 ± 1.0 ka (Czaplewski and Mead et al. 1999). Furlong (1943) investigated a large series of Stockoceros from San Josecito Cave, Mexico, and also utilized material from Shelter Cave and S. onusrosagris material from Papago Springs Cave. His conclusion (p. 5) was "[A] review of the present series demonstrates a gradation from the type of horn core found in the young to those with more divergent tines found in older animals. Thus there appears to be no valid reason for regarding T. conklingi and T. onusrosagris as specifically distinct." Skinner (1942) published a major study of S. onusrosagris from Papago Springs Cave based on remains of at least 125 individuals.
Suites of measurements of the extinct pronghorns are hard to come by in the literature, making it difficult to determine the likelihood that named forms are separate species (Furlong 1943). Stock (1930, 1932) gives ranges for S. conklingi based on a limited number of elements and a few measurements of Capromeryx minor on the basis of one to two specimens. Furlong (1943) gives a range of metacarpal length for S. conklingi from San Josecito Cave, Mexico, based on 92 adult metacarpals and resulting in a range of variation that overlaps the few available measurements of S. onusrosagris. The difference from Stock's small sample and Furlong's large sample is the difference between 147.5-184.6 and 150.0-202.0. Skinner (1942) gave minima, maxima, and means for six limb measurements (unfortunately, other measurements were presented as ratios rather than as raw data). In his sample of metacarpals, the size range of 123 specimens was from 179.3 to 202.5 mm (however, judging from his diagrams, his measurements may be a few millimeters shorter than those based on greatest length). A few other measurements are available in the literature (Colbert and Chaffee 1939, Frick 1937, Roosevelt and Burden 1934).
Despite average differences between the Papago Springs Cave material and material identified as S. conklingi, there is overlap in size of post-cranial material. The horn cores of the fossil taxa appear to show little or no difference between the sexes (Colbert and Chaffee 1939, Furlong 1943). Based on Skinner's measurements, the Papago Springs Cave populations do appear to average larger than the limited sample of S. conklingi. However, it seems plausible that although little sexual difference is displayed in horn core morphology of the fossil species, post-cranial size differences may have occurred between the sexes similar to that seen in the modern Pronghorn, accounting for the observed variations in size. Skinner (1942), noted that there were skulls that might represent females, but also (p. 201) suggested all might be male:
The entire cave collection may represent a seasonal phenomenon, in which only the males frequented the locality. This is not improbable, for the fauna shows a pronounced lack of immature individuals who would have been with the females after fawning time. Male and female Antilocapra are known to separate into sexual herds in winter and early spring previous to the fawning period. The herds again mingle in late summer and early fall.
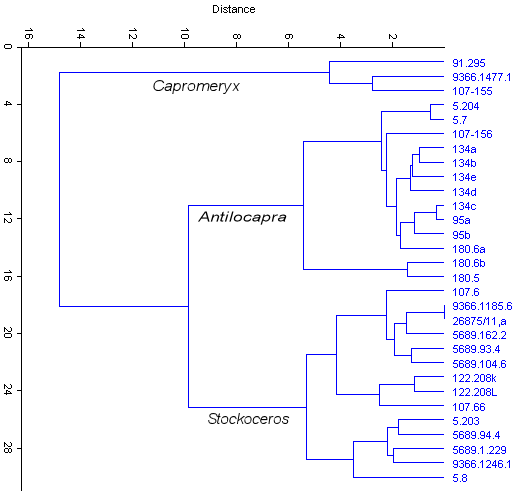 Cluster analysis of the few measurements available
to me of S. conklingi generally show two clusters that arguably, though certainly not
conclusively, could represent size differences between males and females. Fig. 2 is a cluster
analysis (Euclidean distance) based on four measurements of phalanx I (total length, proximal
anterior-posterior depth, proximal width, and mid-shaft width). Specimens interpreted as S.
conklingi divide into two groups at about the -5.5 similarity level, with U-Bar Cave specimens
equally split between the clusters. Similarly, Fig. 3 shows the Shelter Cave specimens dividing
into two separate groupings.
Cluster analysis of the few measurements available
to me of S. conklingi generally show two clusters that arguably, though certainly not
conclusively, could represent size differences between males and females. Fig. 2 is a cluster
analysis (Euclidean distance) based on four measurements of phalanx I (total length, proximal
anterior-posterior depth, proximal width, and mid-shaft width). Specimens interpreted as S.
conklingi divide into two groups at about the -5.5 similarity level, with U-Bar Cave specimens
equally split between the clusters. Similarly, Fig. 3 shows the Shelter Cave specimens dividing
into two separate groupings.
In Fig. 2, all specimens on the Antilocapra branch are modern except those from UTEP Loc. 5, with all modern specimens correctly placed (UTEP 180 is an archaeological site in northern New Mexico). UTEP 134 phalanges are from the same male individual and UTEP 95 phalanges are from the same female individual.
Fig. 2 (left). Similarity cluster dendrogram based on the first phalanges of Antilocapra americana, Capromeryx sp., and Stockoceros conklingi. Some idea of variability is shown in that the A. americana phalanges labeled 134 are from the same individual.
Although Fig. 2 arguably divides the antilocaprids into three taxa, 5.8 and 5.203 are seen in other types of analyses to be in an area of overlap between Antilocapra and Stockoceros. Thus, though such techniques as cluster analysis are suggestive, the results should be viewed with caution.
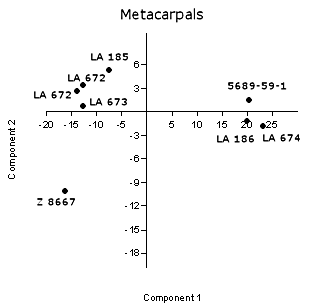 Fig. 3 (right). A principal components plot
applied to antilocaprid metacarpals shows the separation of S. conklingi specimens along the
first axis, primarily a measure of size. Z 8667 is Capromeryx.
Fig. 3 (right). A principal components plot
applied to antilocaprid metacarpals shows the separation of S. conklingi specimens along the
first axis, primarily a measure of size. Z 8667 is Capromeryx.
Another feature to consider is the unlikelihood of two closely related species of ruminants of overlapping size occurring in the same geographic range.
At present, I am taking the case for two species as unproven and, as a result, tentatively follow Furlong (1943) in treating S. onusrosagris as a synonym of S. conklingi.

Fig. 4. From top to bottom: anterior, posterior, and medial views of the right metatarsals of Stockoceros conklingi; at bottom, proximal views of S. conklingi (left) and Antilocapra americana. S. conklingi, UTEP 12-143, Balcony Room site of Dry Cave; modern Pronghorn, UTEP 134. Scale of top three images is 30% of bottom two. Scales in mm.

Fig. 5. Posterior views of the right metacarpals of modern Antilocapra americana (top) and Stockoceros conklingi (UTEP 75-11, Dark Canyon Cave).
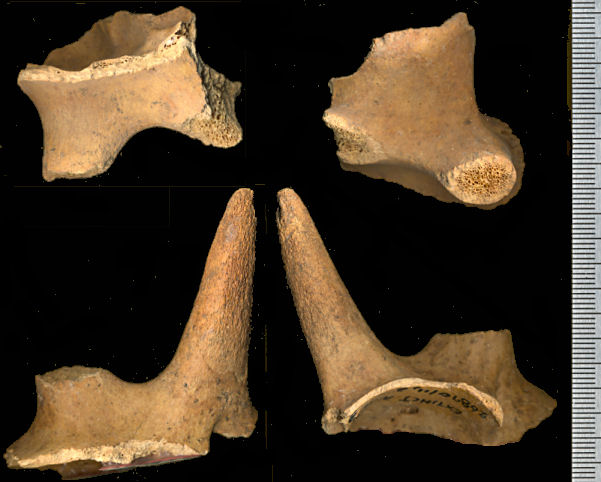
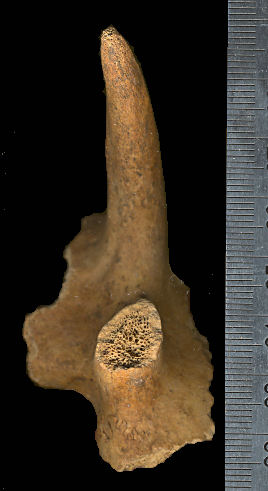 Fig. 6 (left). Right (top) and left partial
frontals with horn core bases and, on left frontal, the anterior horn core (two views of each).
Stockoceros conklingi, U-Bar Cave (5689-24-4a). The specimens are from the original
archaeological excavations. Scale in mm.
Fig. 6 (left). Right (top) and left partial
frontals with horn core bases and, on left frontal, the anterior horn core (two views of each).
Stockoceros conklingi, U-Bar Cave (5689-24-4a). The specimens are from the original
archaeological excavations. Scale in mm.
Fig. 7 (right). Dorsal view of left frontal of Stockoceros conklingi (specimen pictured at bottom of Fig. 6). The shelf on the left is the top of the orbit; a fragment of the parietal is visible at the posterior of the specimen. Scale in mm.
Tebedge (1988) recorded Tetrameryx sp. from Dark Canyon Cave based on a skull fragment with one horn core and the base of the other. In text, the horn core is given as a left posterior; the caption of the figured specimen states it is the right anterior. The latter appears to be correct based on the illustration. The specimen is recorded here as S. conklingi.
Sites. (Includes material listed in the regional literature as Stockoceros sp., Stockoceros conklingi, S. onusrosagris, and as Tetrameryx).
Pleistocene: Ceremonial Cave (Sayles 1935).
Medial Irvingtonian: Slaughter Canyon Cave (Morgan and Harris 2015: cf.).
Rancholabrean: Papago Springs Cave (Skinner 1942).
Early Wisconsin: Papago Springs Cave (Czaplewski and Mead et al. 1999).
Wisconsin: Big Manhole Cave (Harris 1993c: cf.).
Early/Mid Wisconsin: Lost Valley (UTEP).
Mid Wisconsin: Papago Springs Cave (Czaplewski and Mead et al. 1999); Pendejo Cave (Harris 2003); Térapa (Mead et al. 2006); U-Bar Cave (Harris 1987).
Mid/Late Wisconsin: Dark Canyon Cave (Tebedge 1988); Shelter Cave (Stock 1930, 1932).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Balcony Room (UTEP; formerly identified as "Cf. Capromeryx sp": Harris 1993c); Cueva Quebrada (Lundelius 1984: Stockoceros sp.); Pendejo Cave (Harris 2003); Muskox Cave (Logan 1981); Mystery Light Cave (this volume: cf.); U-Bar Cave 13-14 ka (Harris 1989); U-Bar Cave 14-15 ka (Harris 1989); U-Bar Cave 15-18 ka (Harris 1989); U-Bar Cave 18-20 (Harris 1989: cf.); Ventana Cave (Colbert 1950: cf.).
Late Wisconsin/Holocene: Burnet Cave (Schultz and Howard 1935); Pendejo Cave (Harris 2003).
Literature.
Colbert 1950; Colbert and Chaffee 1939; Conkling 1932; Czaplewski and Mead et al. 1999; Frick 1937; Furlong 1943; Harris 1987, 1989, 1993c, 2003; Kurtén and Anderson 1980; Logan 1981; Lundelius 1984; Mead et al. 2006; Morgan and Harris 2015; Roosevelt and Burden 1934; Sayles 1935; Schultz and Howard 1935; Skinner 1942; Stock 1930, 1932; Tebedge 1988.
Last Update: 4 Feb 2016