Class Mammalia
Order Rodentia
Family Cricetidae
Subfamily Sigmodontinae
Sigmodon arizonae—Arizona Cotton Rat // Sigmodon curtisi—Arizona Cotton Rat // Sigmodon hispidus—Hispid Cotton Rat // Sigmodon medius—Intermediate Cotton Rat // Sigmodon minor—Dwarf Cotton Rat //Sigmodon ochrognathus—Yellow-nosed Cotton Rat
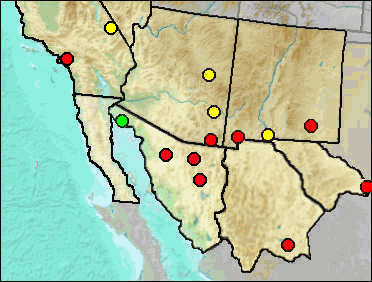
Cotton rats often are considered convergent in ecological niche to the voles, but adapted to warm climatic conditions rather than the cool climates where Microtus centers. Like voles, cotton rats usually are associated with grasses, form runways, and may be active diurnally. Although with some morphological characteristics similar to those of voles, such as short legs and tail, lax fur, and small ears, the dentition differs greatly. The name Sigmodon refers to the more or less "S" shape formed by the cusps, especially those of the third molar.
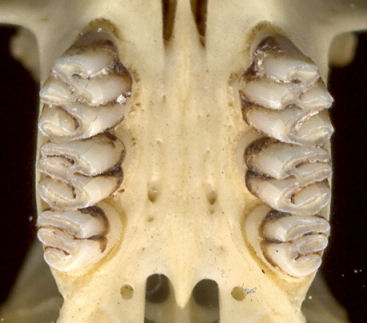

Fig. 1 (left). Palate of Sigmodon hispidus illustrating dental and palatal characteristics. Fig. 2 (right). Right lower toothrow of Sigmodon hispidus showing dental characteristics.
Four species occur now in our region. Sigmodon hispidus is centered to the east of our region, but occupies the Trans-Pecos and the southeastern third of New Mexico west to about the Arizona state line; it is replaced to the west by S. arizonensis, with S. fulviventer occurring in the southwestern part of New Mexico and with an apparently isolated population in the New Mexican north-central Rio Grande Valley. Sigmodon ochrognathus occurs in the mountains of the Trans-Pecos, extreme southern New Mexico and in scattered areas in Arizona. All species have an extensive present to the south in Mexico.
Sites.
Late Blancan: La Union (Morgan and Lucas 2003); San Simon Fauna (Morgan and White 2005); Snowflake (Lindsay and Tessman 1974); Tecopa Lake Beds (Woodburne and Whistler 1991).
Irvingtonian: El Golfo (Croxen et al. 2007).
?Irvingtonian/Rancholabrean: Emery Borrow Pit (Jefferson 1991b).
Rancholabrean: La Playa (White et al. 2010); Térapa (White et al. 2010).
Sangamon: La Brisca (Van Devender et al. 1985).
Early Mid or Early Wisconsin: Lost Valley (Harris 1993c).
Mid/Late Wisconsin/Holocene: Jimenez Cave (Messing 1986).
Late Wisconsin: Algerita Blossom Cave (Harris 1993c); Bonfire Shelter (Frank 1968); Harris' Pocket (UTEP); Human Corridor (Harris 1993c); Lehner Ranch (Lindsay and Tessman 1974); Murray Springs (Lindsay and Tessman 1974).
Late Wisconsin/Holocene: Beyond Bison Chamber (UTEP); Howell's Ridge Cave (Harris 1993c).
Literature. Croxen et al. 2007; Frank 1968; Harris 1993c; Messing 1986; Morgan and Lucas 2003; Van Devender et al. 1985; White et al. 2010); Woodburne and Whistler 1991.
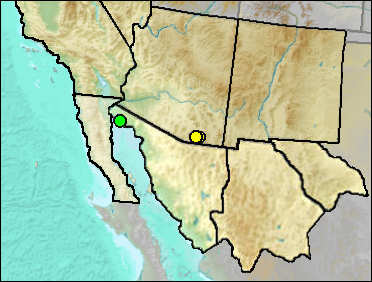
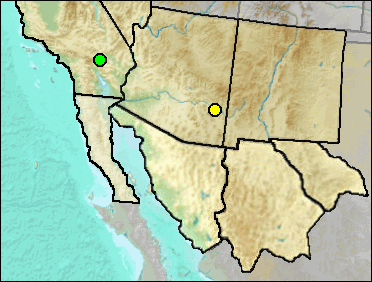
The Arizona Cotton Rat occurs very marginally in southwestern New Mexico and in southeastern Arizona on into Mexico. It is very similar to the Hispid Cotton Rat.
Sites.
Mid Wisconsin: Papago Springs Cave (Czaplewski and Mead et al. 1999: cf. sp.).
Late Wisconsin/Holocene: Deadman Cave (Mead et al. 1984: cf.).
Literature. Czaplewski and Mead et al. 1999
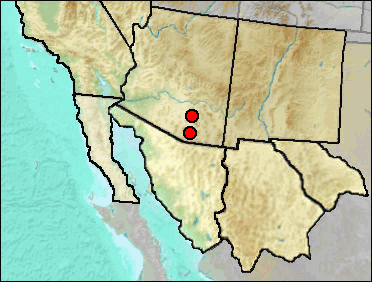
This cotton rat approaches modern species of Sigmodon in terms of hypsodonty, but has three roots to m1 rather the four found in the living species of the United States (Kurtén and Anderson 1980).
Sites.
Late Blancan: California Wash (Johnson et al. 1975); Curtis Ranch (Lindsay 1984).
Irvingtonian: El Golfo (Croxen et al. 2007).
Literature. Croxen et al. 2007; Johnson et al. 1975; Lindsay 1984.
 Sigmodon hispidus is
the common species in the region (Fig. 1). At one time, the species was thought to
continue westward in Arizona; more recently, it was learned that the adjacent
populations in Arizona and overlapping slightly into New Mexico represented a cryptic
species, Sigmodon arizonae. It is quite possible that U-Bar Cave specimens
represent that species rather than S. hispidus; characteristics that could
separate the two are unknown for the fossil specimens available.
Sigmodon hispidus is
the common species in the region (Fig. 1). At one time, the species was thought to
continue westward in Arizona; more recently, it was learned that the adjacent
populations in Arizona and overlapping slightly into New Mexico represented a cryptic
species, Sigmodon arizonae. It is quite possible that U-Bar Cave specimens
represent that species rather than S. hispidus; characteristics that could
separate the two are unknown for the fossil specimens available.
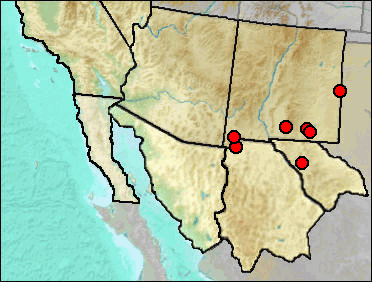
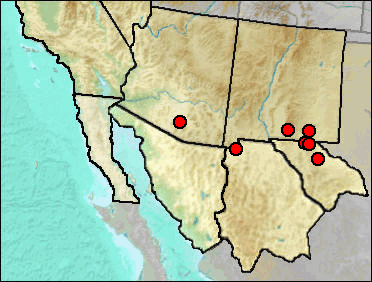
Fig. 1. Approximate current geographic range of the Hispid Cotton Rat in our region.
The Holocene record from Atlatl Cave in San Juan County, NM, is listed (but not mapped) because it indicates a major expansion of range, presumably during a time when grasslands allowed passage across the continental divide separating the Rio Grande drainage from that of the San Juan River.

Fig. 2. Lateral view of left dentary and ventral view of Sigmodon hispidus skull (UTEP mammal collection 1358). Scale in mm.
Sites.
Mid Wisconsin: Pendejo Cave (Harris 2003); U-Bar Cave (Harris 1987).
Mid/Late Wisconsin: Dark Canyon Cave (Tebedge 1988)
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP).
Late Wisconsin: Blackwater Loc. No. 1 (Slaughter 1975); TT II (Harris 1993c: cf.).
Late Wisconsin/Holocene: Balcony Room (Harris 1993c); Howell's Ridge Cave (UTEP); Pendejo Cave (UTEP).
Middle Holocene: Atlatl Cave (Gillespie 1985).
Literature. Dalquest and Stangl 1984b; Gillespie 1985; Harris 1987, 1993c, 2003; Slaughter 1975; Tebedge 1988.
Gidley (1922:126) described S. medius from the Benson beds of Arizona:
Description.-Length of cheek-tooth series 5.5 millimeters; somewhat smaller than the living species S. sanctae martae, with which it seems to compare more nearly than with other living forms. Its resemblances are noted in the form of the anterior lobe of the first cheek tooth, in which the anterior internal reentrant valley considerably exceeds the opposing outer one in length; in the decided angulation of the posterior portion of the inner wall of the last molar; and in the relatively heavy symphysis and considerable depth of jaw. It differs from this and other living species in having the cheek-tooth crowns slightly less hypsodont, the reentrant valleys more open, and the lophs correspondingly narrower, while there is present in the last molar a small but distinct reentrant notch on its inner side opposite the posterior reentrant valley.
The two upper cheek teeth show the same character of the more open valleys observed in the lower cheek teeth. Another difference noted is the form of the external reentrant valleys of these teeth, which clearly distinguishes them from the corresponding teeth of living species. The external reentrant valleys, instead of being straight, open directly outward, while their inner portions bend backward, forming a decided angle in their posterior enamel walls. In the living species these valleys run obliquely forward and outward in a nearly straight or but slightly curved line.
Morgan et al. (2011) listed its presence in the Caballo Local Fauna in their Table 1, but their discussion of the taxon implies it does not occur in that portion of the Blancan, and they do not list it among the referred specimens.
Many authors consider S. medius to be a synonym of S. minor. However, Morgan et al. (2011) recognized these as separate species, the latter being significantly smaller than the former. This is the approach followed here.
Sites.
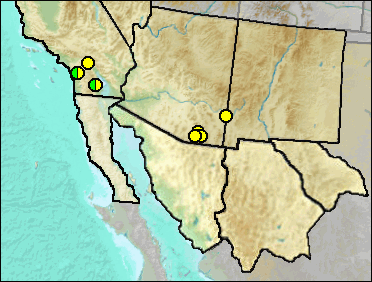
Late Blancan: 111 Ranch (Morgan and White 2005).
Early Irvingtonian: Gypsum Ridge (Wagner and Prothero 2001).
Literature. Morgan and White 2005; Morgan et al. 2011; Wagner and Prothero 2001.
As indicated the S. medius account, Morgan et al. (2011) recognized S. minor as separate from S. medius; they also considered it an important biostratigraphic marker of the latest Blancan.
Gidley (1922:125) described S. minor in the same paper as S. medius:
Description.-Length of cheek-tooth series 4.7 millimeters; smaller than S. medius and decidedly smaller than any of the living species examined. This species is distinguished from the living forms by the same characters observed in S. medius, except that the external reentrant valleys of the upper molars (as shown by specimen No. 10513) are of the normal modern type. S. minor differs from S. medius only in its smaller size, having a relatively narrower anterior lower cheek tooth, in which the anterior lobe is relatively smaller, with the adjacent reentrant valleys nearly equal in length. The reentrant valleys of the upper cheek teeth are oblique to the same degree and of about the same proportionate depth, but the molar crowns of both the upper and lower series are less hypsodont.
Sites.
Late Blancan: California Wash (Johnson et al. 1975); Curtis Ranch (Lindsay 1984); Jack Rabbit Trail, San Timoteo Badlands (Albright 2000); Virden (Morgan et al. 2011: cf.).
Late Blancan/Early Irvingtonian: Elsinore: Mimomys (Pajak et al. 1996).
Irvingtonian: Anza-Borrego (Murray 2008).
Literature. Albright 2000; Johnson et al. 1975; Lindsay 1984; Morgan et al. 2011; Murray 2008; Pajak et al. 1996.
The Yellow-nosed Cotton Rat generally has been considered as a higher-elevation rodent than S. hispidus. In our region, it occurs in the mountains of the New Mexican bootheel, and in Texas in a number of localities in the Trans-Pecos, including in recent years in non-montane habitats (Schmidly 2004). Hollander et al. (1990) reported it from the Guadalupe Mountains, some 80 miles north of its earlier recognized range, putting it in striking distance of several important Pleistocene fossil sites. It has been reported from Holocene deposits (ca. 3580 BP) of Big Manhole Cave (Lear and Harris 2007) and Pratt Cave (Stangl and Dalquest 1991), though there is some question as to whether Pleistocene material is mixed into the Holocene material at the latter site (see the Pratt Cave account).
 Stangl
and Dalquest (1990) investigated dental differences between S. hispidus and
S. ochrognathus. They found that the upper toothrow is smaller in S.
ochrognathus at comparable degrees of wear, and especially pronounced with M1.
Their measurements of the occlusal surfaces of M1 (in mm) gave a mean of 2.29 ± 0.10
(s.d.) for S. hispidus compared to 2.02 ± 0.10 for S. ochrognathus, and
for width, respectively, 1.73 ± 0.10 and 1.51 ± 0.10. Extremes, however, did
overlap.
Stangl
and Dalquest (1990) investigated dental differences between S. hispidus and
S. ochrognathus. They found that the upper toothrow is smaller in S.
ochrognathus at comparable degrees of wear, and especially pronounced with M1.
Their measurements of the occlusal surfaces of M1 (in mm) gave a mean of 2.29 ± 0.10
(s.d.) for S. hispidus compared to 2.02 ± 0.10 for S. ochrognathus, and
for width, respectively, 1.73 ± 0.10 and 1.51 ± 0.10. Extremes, however, did
overlap.
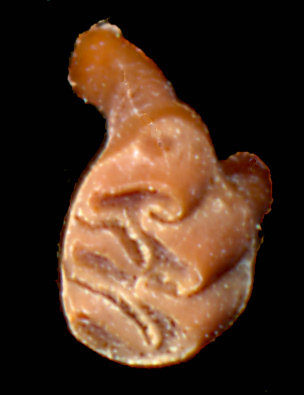
Fig. 1. Upper right first molar of fossil Sigmodon ochrognathus from Dust Cave.
Another character noted by Stangl and Dalquest concerned the anterocone of M1; they found it compressed or flattened anteriorly in 47 of 50 S. ochrognathus as compared to a usually convex anterocone in S. hispidus. The latter, however, did show compression in 12 out of 50 specimens of S. hispidus. The anterior of the anterocone, viewed laterally, was said to form an even slope from the alveolus to the occlusal surface in S. hispidus, but to be recessed posteriorly in S. ochrognathus.
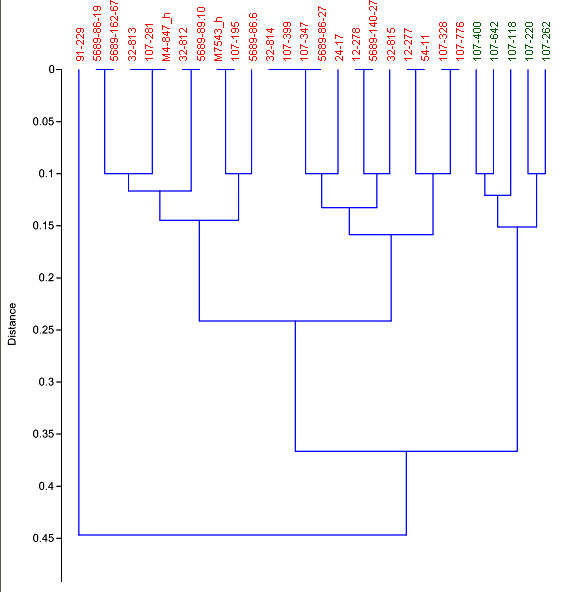
Fossil specimens of m1 interpreted as S. ochrognathus are small, and when width and length are taken together, appear to separate them from those of S. hispidus (Fig. 2). Measurements follow Ruez (2011) except that m1 length is taken from the lingual side, parallel to the occlusal surface, from the anterior junction of enamel with dentine.
Fig. 2. Cluster diagram (paired group, Euclidean distance) based on length by width of m1. Red is interpreted as Sigmodon hispidus and green as S. ochrognathus. The outlier (91-229) has an m1 as narrow as the latter species, but is longer than any other m1 in the sample.

Fig. 3. Sigmodon ochrognathus. Photograph by Roger W. Barbour, courtesy of the Smithsonian National Museum of Natural History.
Sites.
Early/Early-Mid Wisconsin: Lost Valley (UTEP).
Mid/Late Wisconsin/Holocene: Sierra Diablo Cave (UTEP cf.)
Late Wisconsin: Dust Cave (Harris and Hearst 2012); Pendejo Cave (UTEP); U-Bar Cave 15-18 ka (Harris 1989).
Late Wisconsin/Holocene: Fowlkes Cave (Stangl and Dalquest 1991); Pendejo Cave (UTEP); Pratt Cave (Stangl and Dalquest 1991); Wolcott Peak (Mead et al. 1983).
Literature. Harris 1989; Harris and Hearst 2012; Hollander et al. 1990; Lear and Harris 2007; Mead et al. 1983; Ruez 2011; Schmidly 2004; Stangl and Dalquest 1991.
Last Update: 17 Jul 2014